XiaoMi-AI文件搜索系统
World File Search System存储


UVA-dare(数字学术存储库)
作为一个相对年轻的研究领域,定量风险管理将定量技术应用于风险管理学科。数学模型和技术有助于量化金融系统复杂动态的风险。这些模型的范围和应用是巨大的;例如,它们包括银行业中的信用风险投资组合模型,保险部门的破坏理论模型,养老金部门的资产责任管理模型以及在所有金融部门应用的气候风险方法论。量化风险管理是一项迅速发展的纪律,也是对金融业的持续挑战,不仅是由经济动荡,政治不稳定或共同发展等外部发展所助长的,而且还受到新法规的挑战。除了对金融机构当前风险和财务状况的深入分析之外,前瞻性的定量风险管理是了解未来风险及其对这些机构的影响的关键,以便准备或适应不需要的事件。

弗洛尔存储库istituzionaledell'universitàDeglistudi ...
其中e n =(0,。。。,1)∈Rn,我们设置:q(x)=(1 + | x | 2) / 2。< / div>已经为Alexandrov的Soap Bubble定理获得了此类的整体身份,以及该注释的作者经典的Serrin问题(请参阅[14、15、16、20])。在那些情况下,X Q在身份中的作用是由身份字段rn∋x7→x扮演的。请注意,在单位球体上,x q是s上的 - e n的投影。在[8]中,证明,如果您满意(1.1) - (1.2),则(1.5)的左侧必须为零。自从x n> 0中的x n> 0在ωby[8,命题2.3]中的u <0中,(1.5)左侧的牙套中的功能必须在ω上消失相同,因为cauchy-schwarz始终是非负性的。作为一种副产品,一个人必须是u必须是球形对称的二次多项式,如[14]中所述。因此,σ必须是球的一部分,因为σ上的u = 0。ω的凸耳形状很容易随机。现在,观察到(1.5)很明显其右侧(因此其左侧)是无效的,如果(1.2)持有(1.2)。但是,(1.5)至少有两个原因提供了更多信息。一个是,在较弱的假设(1.2)的右侧是非阳性的较弱的假设下,郭和夏的刚性结果只能获得。第二个也是更重要的原因是该身份提供了定量信息。实际上,如果我们知道uν在某种整体规范中与r偏离R,那么(1.5)的左侧积分很小。现在,请注意,如果我们将二次多项式视为
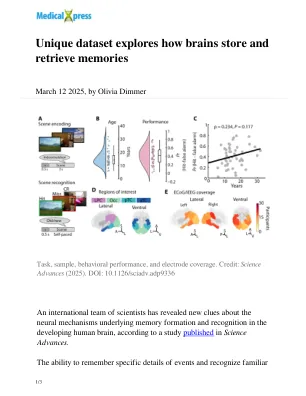
唯一数据集探讨了大脑如何存储和检索记忆
虽然科学家已经能够研究参与记忆形成和检索的大脑部分,但这些过程是如何由大脑的各个部分实施的。虽然科学家已经能够研究参与记忆形成和检索的大脑部分,但这些过程是如何由大脑的各个部分实施的。

2025年3月11日,康涅狄格州太阳能和存储的评论...
太阳能光伏系统。Connssa是该州的住宅太阳能安装商和商业太阳能开发商的业务组,雇用了2,000多名居民,他们在康涅狄格州提供,出售,设计,工程和安装太阳能电动(光伏)能源产品。已安装在100,000多个康涅狄格州的住宅和商业场所上。Connssa不支持HB 6052。我们同意HB 6052的某些部分。直言不讳,我们已经公开同意三(3)个不同但类似的家庭太阳能消费者保护法案中的一部分。在2021年,SB 951关于住宅太阳能光伏系统招标的法案。康涅狄格州太阳能行业愿意与帐单支持者会面,并在第二年退还更好的账单。我们没有接受我们在书面证词中提供的会面的提议。在2022年是No.5326关于住宅太阳能光伏系统招标和涉及太阳能的房地产交易的法案。再次,康涅狄格州太阳能安装人员提出与帐单支持者会面。无响应。在2024年,SB 297关于太阳能公司客户的消费者保护的一项法案。它创建了一个由16名成员组成的工作组,以研究改善客户太阳能光伏系统销售的方法。工作队将在2025年1月1日之前向立法机关报告其调查结果和建议。现在(HB 6052)再次发生。几位太阳能行业志愿者提出要担任工作队,并由州长拉蒙特(Lamont)的工作人员和核心小组领导人分别审查,并被任命为从未举行过会议的工作队。令人失望的是,在前三(3)次尝试通过太阳能销售消费者保护费用太阳能安装人员被排除在初次讨论之外。自从我们在2021年提供帮助以来,我们对太阳能销售消费者保护政策的看法没有改变。我们继续分享一些相同的合理立法政策思想。实际上,我们的成员已经与许多其他州合作,在该领域发展了最佳实践。我们渴望分享其他州发现的坚实政策。我们当然反对冗余政策,这些政策为销售集团和市政当局和强烈反对法规创造了额外的工作,这些法规将以太阳能行业的规则和罚款来挑选出HB 6052中的规则和惩罚,而HB 6052中并未在所有房屋建筑销售中同样应用。

第1章 - UVA -DARE(数字学术存储库)
如今,通过各种高通量技术的开发,可以很好地分析真核基因组的线性维度,从而可以进行基因组范围的方法。因此,他们的序列几乎没有谜,更容易质疑他们的进化和越来越多的研究旨在绘制其动态表观基因症状。这一进展引起了新的挑战,即使基因组重新恢复其三维核框架,以检查基因组的主要功能与相互相间细胞核的结构之间的相互作用,从而破译了核结构与功能之间的关系。因此,对核室有新的兴趣,其中一些描述了大约两个世纪前和3D核结构。因此,在动物和植物细胞中都在积极研究了相间细胞核的特殊复杂性,其有序结构以及该细胞器的动力学。已经了解了细胞核的组成和精细结构,以及其各种功能隔室的形成机理和动力学的机理。对染色质和其他核室之间的结构和功能相互作用有了更好的了解。这些研究伴随着特定的3D方法和工具的开发,例如3D成像和建模以及捕获染色体构象的方法。然而,关于植物中的染色质动力学还有很多尚待了解。已经发表了许多关于核组织各个方面的评论(De Wit and de Laat 2012; Dekker等,2013; Delgado等,2010; Dion and Gasser 2013; Rajapakse and Groudine 2011; Taddei and Gasser 2012; Towbin等,2012; Towbin等人,2013年)。在这篇综述中,我们总结了我们当前对模型植物拟南芥中相间核核区室的知识,并特别强调了异染色质。的确,这个隔室是高度塑料的,表现出大规模的重组并有助于基因组组织,而在细胞核尺度上的白染色质动力学几乎没有研究。我们还讨论了3D建模和定量技术,用于分析相互核的体系结构,这些核的结构仍处于thaliana的起步阶段。

PoliteCnico DI Torino存储库Istituzionale
反应性氧化物(ROS)对活细胞生存能力和增殖的影响很多。由于它们与不同类型的生物分子反应的能力,ROS参与了许多细胞功能1。维持氧化还原稳态的能力至关重要,失衡会导致各种可能的疾病。可以利用受控的ROS产生以产生细胞中的氧化应激,导致细胞死亡,目的是开发用于抗癌治疗的药物和无药物治疗工具。氨基丙基官能化的ZnO NC(ZnO-NH 2 NC)被证明可以使用已批准的医疗设备Lipozero G39刺激超声(US)时,能够以可调且可重复的方式产生ROS。羟基自由基的产生是美国暴露下惯性空化的结果。

可再生能源分区,景观特征评估,电池电气存储系统和生物多样性
Lixnaw Wind Awine Group,Luachra Wind Awarness Group,Tullamore Action Group,North Kerry Wind Turbine


UOC存储库
[1] M. V. Chao,“神经营养蛋白及其受体:许多信号通路的收敛点”,Nat。修订版Neurosci。,卷。4,不。4,pp。299–309,2003。[2] M. Bothwell,“ NGF,BDNF,NT3和NT4”,在神经营养因素中。实验药理学手册,施普林格,柏林,海德堡,2014年。[3] R. Levi-Montalcini,H。Meyer和V. Hamburger,“体外实验对小鼠肉瘤180和37对雏鸡胚胎的感觉和交感神经系统的影响,”癌症Res。,1954年。[4] R. Levi-Montalcini,“ 35年后的神经生长因子”,科学(80-。)。,1987。[5] Y.A. Barde,D。Edgar和H. Thoenen,“哺乳动物大脑的新神经营养因子的纯化”,Embo J.,1982。[6] K. R. Jones和L. F. Reichardt,“人类基因的分子克隆,该基因是神经生长因子家族的成员。”natl。学院。SCI。 U. S. A.,1990。 [7] P. C. Maisonpierre等。 ,“神经营养蛋白3:与NGF和BDNF有关的神经营养因子”,科学(80-。 )。 ,1990。 [8] A. Hohn,J。Leibrock,K。Bailey和Y. A. Barde,“神经生长因子/脑源性神经营养因子家族的新成员的识别和表征”,自然,1990年。 [9] A. Rosenthal等。 ,“新型人类神经营养因子的主要结构和生物学活性”,Neuron,1990。 [10] N. Y. IP等。 natl。 学院。 SCI。SCI。U. S. A.,1990。[7] P. C. Maisonpierre等。,“神经营养蛋白3:与NGF和BDNF有关的神经营养因子”,科学(80-。)。,1990。[8] A. Hohn,J。Leibrock,K。Bailey和Y.A. Barde,“神经生长因子/脑源性神经营养因子家族的新成员的识别和表征”,自然,1990年。[9] A. Rosenthal等。,“新型人类神经营养因子的主要结构和生物学活性”,Neuron,1990。[10] N. Y. IP等。natl。学院。SCI。SCI。,“哺乳动物神经营养蛋白4:结构,染色体定位,组织分布和受体特异性。”U. S. A.,1992。[11] R. Gotz等。,“ Neurotrophin-6是神经生长因子家族的新成员”,自然,1994年。[12] K. O. Lai,W。Y. Fu,F。C. F. Ip和N. Y.单元格。Neurosci。,1998。[13] M. A. Bothwell和E. M. Shopter,“β神经生长因子的离解平衡常数”,J Biol Chem,1977。[14] C. Radziejewski,R。C。Robinson,P。S。S. Distefano和J. W. Taylor,“脑源性神经营养因子和神经营养因子和神经营养蛋白3。的二聚体结构和构象稳定性,” Biiochemistry,1992。[15] M. J. Butte,P。K。Hwang,W。C。Mobley和R. J. Fletterick,“ Neurotrophin-3同二聚体的晶体结构显示出不同的区域用于结合其受体,” 1998年。[16] N.[17] R. C. Robinson等。,“神经营养蛋白4同二聚体的结构和脑衍生的神经营养因子/神经营养蛋白4异二聚体揭示了一个常见的TRK结合位点,”蛋白质SCI。,2008。[18] K. K. Teng,S。Felice,T。Kim和B. L. Hempstead,“了解胸部营养蛋白的作用:最近的进步和挑战”,发展性神经生物学。2010。401–3,1992。:ebsCohost,” Annu。修订版Neurosci。[19] G. CM,“通过生理活性调节脑神经营养蛋白表达。”趋势Pharmacol Sci,pp。[20] S. D. Skaper,“神经营养因素:概述”,《分子生物学方法》,2018年。[21] A. K. McAllister,L。C。Katz和D. C. Lo,“神经营养蛋白和突触可塑性。,1999。[22] S. Pezet和S. B. McMahon,“神经营养蛋白:疼痛的介体和调节剂”,Annu。修订版Neurosci。,2006年。[23] D. R. Kaplan,B。L。Hempstead,D。Martin-Zanca,M。V。Chao和L. F. Parada,“ TRK原型癌基产品:神经生长因子的信号传递受体,”科学(80-。)。,1991。[24] R. Klein等。,“ TRKB酪氨酸蛋白激酶是脑源性神经营养因子