XiaoMi-AI文件搜索系统
World File Search System1A1

英国温室气体清单,1990 年至 2019 年 - UK-AIR
IPCC 类别 来源 描述 1 能源 1A 燃料燃烧活动 1A1 能源产业 1A1a 公共电力和热力生产 1A1b 石油精炼 1A1c 固体燃料制造及其他能源产业 1A2 制造业和建筑业 1A2a 钢铁 1A2b 有色金属 1A2c 化学品 1A2d 纸浆、造纸和印刷 1A2e 食品加工、饮料和烟草 1A2f 非金属矿物 1A2gvii 制造业和建筑业中的移动燃烧 1A2gviii 制造业和建筑业中的固定燃烧:其他 1A3 运输 1A3ai 国际航空 1A3aii 民航 1A3b 公路运输 1A3c 铁路 1A3d 国内航运 1A3e 其他(待指定) 1A4 其他部门 1A4a 商业/机构燃烧 1A4b 住宅 1A4c 农业/林业/渔业 1A5 其他(未另行说明) 1A5a 其他,固定式(包括军用) 1A5b 其他,移动式(包括军用)

癌症抗药性
癌症是全球死亡的第二大原因。癌症患者的存活取决于疗法的功效和抗药性的发展。癌细胞获取耐药性,包括获得类茎样特征,涉及许多机制。癌症干细胞(CSC)代表了肿瘤进展和耐药性的主要来源。CSC是具有自我更新和体外形成球体能力的癌细胞的亚群。醛脱氢酶1a1(Aldh1a1)是一种胞质酶,参与了来自毒性醛的细胞排毒,属于Aldh家族。高ALDH1A1活性与几种肿瘤的干性表型密切相关,可能导致体内癌症的进展和扩散。 我们通过激活缺氧诱导因子-1α和血管内皮生长因子信号传导的激活来证明ALDH1A1在乳腺癌细胞中肿瘤血管生成中的贡献。 本综述讨论了Aldh1a1参与不同癌症标志的发展,以将其作为癌症治疗的新型推定靶标,以取得更好的预后。 在这里,我们分析了AldH1A1在肿瘤细胞中的摄入干性表型中的参与,肿瘤血管生成和转移的调节以及抗癌耐药性和免疫逃避性的获取。高ALDH1A1活性与几种肿瘤的干性表型密切相关,可能导致体内癌症的进展和扩散。我们通过激活缺氧诱导因子-1α和血管内皮生长因子信号传导的激活来证明ALDH1A1在乳腺癌细胞中肿瘤血管生成中的贡献。本综述讨论了Aldh1a1参与不同癌症标志的发展,以将其作为癌症治疗的新型推定靶标,以取得更好的预后。在这里,我们分析了AldH1A1在肿瘤细胞中的摄入干性表型中的参与,肿瘤血管生成和转移的调节以及抗癌耐药性和免疫逃避性的获取。

基于靶向定量蛋白质组学的小肠药物转运蛋白和代谢酶的个体发育 Kiss, M.; Mbasu, Richard; Nicolai, J
儿童大部分药物为口服给药,但各年龄段儿童小肠药物代谢酶(DME)和药物转运体(DT)的蛋白质丰度信息仍不明确,这阻碍了儿童精准用药。为了探索 DME 和 DT 的年龄相关差异,收集了儿童和成人空肠和回肠手术剩余的肠组织,并通过靶向定量蛋白质组学分析了顶端钠 - 胆汁酸转运蛋白、乳腺癌耐药蛋白(BCRP)、单羧酸转运蛋白 1(MCT1)、多药耐药蛋白 1(MDR1)、多药耐药相关蛋白(MRP)2、MRP3、有机阴离子转运多肽 2B1、有机阳离子转运蛋白 1、肽转运蛋白 1(PEPT1)、CYP2C19、CYP3A4、CYP3A5、UDP 葡萄糖醛酸转移酶(UGT)1A1、UGT1A10 和 UGT2B7。分析了 58 名儿童(48 条回肠、10 条空肠,年龄范围:8 周至 17 岁)和 16 名成人(8 条回肠、8 条空肠)的样本。比较年龄组时,成人回肠中的 BCRP、MDR1、PEPT1 和 UGT1A1 丰度明显高于儿童回肠。空肠 BCRP、MRP2、UGT1A1 和 CYP3A4 丰度在

AAV8-hUGT1A1 与雷帕霉素联合治疗新生儿感染的疗效...
一项使用腺相关病毒血清型 8 (AAV8)-人尿苷二磷酸葡萄糖醛酸转移酶 1A1 (h UGT1A1) 治疗遗传性严重未结合高胆红素血症 (Crigler-Najjar 综合征) 的临床试验正在进行中,但临床前数据表明,由于在生长中的肝脏中肝细胞增殖时转基因表达丧失,该疗法在儿童中的长期疗效不佳。本研究旨在确定在相关动物模型中在什么年龄可以获得长期疗效,以及免疫调节是否允许使用相同的 AAV 载体进行重复治疗。新生、哺乳期和幼年 Ugt1a1 缺乏大鼠接受了临床相关剂量的 AAV8-h UGT1A1 ,并监测血清胆红素水平和血清中的抗 AAV8 中和抗体 (NAbs)。使用雷帕霉素方案研究了预防对载体的免疫反应的可能性,该方案从载体给药前 2 天开始,到给药后 21 天结束,每天进行腹膜内 (ip) 注射。在出生后第 1 天 (P1) 或 P14 接受治疗的大鼠中,12 周后校正 (部分) 消失,而在 P28 注射的大鼠中校正保持稳定。将初始载体给药与免疫抑制方案相结合可防止雌性大鼠诱导 NAbs,从而至少可以部分有效地进行再次给药。再次注射后无法防止 NAbs 的诱导,这表明该策略对于先前存在的抗 AAV NAbs 水平较低的患者无效。

STAT5 在 CD34+/CD38- 干细胞中表达,并作为 Ph 阴性骨髓增生性肿瘤的潜在分子靶点
1 维也纳医科大学路德维希玻尔兹曼血液学和肿瘤学研究所,1090 维也纳,奥地利;daniela.berger@meduniwien.ac.at (DB);daniel.ivanov@meduniwien.ac.at (DI);yueksel.filik@onc.lbg.ac.at (YF);peter.valent@meduniwien.ac.at (PV) 2 维也纳兽医大学小动物内科医院伴侣动物和马匹科/医院,1210 维也纳,奥地利 3 维也纳医科大学内科 I,血液学和止血学分部,1090 维也纳,奥地利;alexandra.keller@meduniwien.ac.at (AK);florian.schur@ist.ac.at (FKMS); christoph.kornauth@meduniwien.ac.at(CK)4 维也纳医科大学实验医学系,1090 维也纳,奥地利;georg.greiner@meduniwien.ac.at(GG);nadine.witzeneder@meduniwien.ac.at(NW);gregor.hoermann@meduniwien.ac.at(GH)5 维也纳兽医大学动物育种和遗传学研究所,1210 维也纳,奥地利;bettina.wingelhofer@manchester.ac.uk(BW);heidi.neubauer@vetmeduni.ac.at(HAN);richard.moriggl@vetmeduni.ac.at(RM)6 INSERM,ERI-12,皮卡第-儒勒凡尔纳大学药学院,80000 亚眠,法国; emmanuel.pecnard@univ-tours.fr (EP); fabrice.gouilleux@univ-tours.fr (FG) 7 分子医学研究中心(CeMM),1090 维也纳,奥地利; harini.nivarthi@meduniwien.ac.at (HN); robert.kralovics@meduniwien.ac.at (RK) 8 维也纳医科大学病理学系,1090 维也纳,奥地利; leonhard.muellauer@meduniwien.ac.at 9 多伦多大学化学系,多伦多,ON M5S 1A1,加拿大; gary.tin@mail.utoronto.ca (GT); ji.park@mail.utoronto.ca (JP); e.dearaujo@mail.utoronto.ca (EDdA); patrick.gunning@utoronto.ca (PTG) 10 CNRS UMR 6239, GICC, Facult é de Mé decine, Universit é François Rabelais, 37020 Tours, France * 通讯地址: emir.hadzijusufovic@meduniwien.ac.at;电话:+ 43-1-40400-49990

全基因组鉴定肌萎缩侧索硬化症的遗传基础
张赛, 1 , 8 Johnathan Cooper-Knock, 2 , 8 Annika K. Weimer, 1 Minyi Shi, 1 Tobias Moll, 2 Jack NG Marshall, 2 Calum Harvey, 2 Helia Ghahremani Nezhad, 2 John Franklin, 2 Cleide dos Santos Souza, 2 Ke Ning, 2 Cheng Wang, 3 Jingjing Li, 3 Allison A. Diliot, 4 Sali Farhan, 4 Eran Elhaik, 5 Iris Pasniceanu, 2 Matthew R. Livesey, 2 Chen Eitan, 6 Eran Hornstein, 6 Kevin P. Kenna, 7 Project MinE ALS 测序联盟, Jan H. Veldink, 7 Laura Ferraiuolo, 2 Pamela J. Shaw, 2 和 Michael P. Snyder 1 , 9 , * 1 遗传学系中心斯坦福大学医学院基因组学和个性化医学系,斯坦福,CA 94305,美国 2 谢菲尔德大学谢菲尔德转化神经科学研究所,谢菲尔德,S10 2HQ,英国 3 伊莱和埃迪丝布罗德再生医学和干细胞研究中心、巴卡尔计算健康科学研究所、帕克癌症免疫治疗研究所和加州大学旧金山分校医学院神经病学系,旧金山,CA 94143,美国 4 麦吉尔大学蒙特利尔神经病学研究所神经病学和神经外科系,蒙特利尔,QC H3A 1A1,加拿大 5 隆德大学生物系,隆德 223 62,瑞典 6 魏茨曼科学研究所分子遗传学系,雷霍沃特 7610001,以色列 7 大学医学中心鲁道夫马格努斯脑中心神经病学系乌得勒支,乌得勒支 3584 CX,荷兰 8 这些作者贡献相同 9 主要联系人 *通信地址:mpsnyder@stanford.edu https://doi.org/10.1016/j.neuron.2021.12.019

银河合并挑战:基于机器学习的检测方法之间的比较研究
。加巴。计算机科学,UL。。 08544,美国墨西哥蒙丹,88025 34055,韩国10 St. Cherry,Tucson,Tucson,AZ,美国物理与天文学大学3009,NA 6009,15

恢复期血清和疫苗接种血清对 SARS-CoV-2 祖先变体、β、δ 和 omicron 变体临床分离株的免疫原性
康复和接种血清的免疫原性针对临床造成的临床隔离,祖先SARS-COV-2,Beta,三角洲和Omicron变体2 3 Arinjay Banerjee 1,2,3,*,Jocelyne lew 1,liw 1,andrea kroeker 1,andrea kroeker 1,andrea kroeker 1,kaushal baid 1,kaush baid 1,patrick aftich ahah nirm nirm nirm nirm Ryan McDonald 8,Amanda Lang 8.9,5 Volker Gerdts 1.2,Sharon E. Straus 10.11,Lois Gilbert 12,Angel Xinliu li 12,Mohammad 6 Mozafarihasjin 12,Sharon Walmsley 13,Anne-Claude 13,Anne-Claude 12,14,Jeffrey L. Wrana 12,14,Tony 7,tony 7,盖尔7,12,15;加拿大萨斯喀彻温省萨斯卡通 S7N 11 5E3 12 2 萨斯喀彻温大学兽医微生物学系;萨斯喀彻温省萨斯卡通 S7N 5B4,13 加拿大 14 3 滑铁卢大学生物系;加拿大安大略省滑铁卢 N2L 3G1 15 4 共享医院实验室;加拿大安大略省多伦多 M4N 3M5 16 5 Sunnybrook 研究所;加拿大安大略省多伦多 M4N 3M5 17 6 达尔豪斯大学计算机科学学院;加拿大新斯科舍省哈利法克斯 B3H 4R2 18 7 多伦多大学实验室医学和病理生物学系;加拿大安大略省多伦多 19 M5S 1A8 20 8 萨斯喀彻温省卫生局 Roy Romanow 省实验室;里贾纳,SK,S4S 0A4,21 加拿大 22 9 萨斯喀彻温大学医学院;加拿大萨斯喀彻温省萨斯卡通 S7N 5E5 23 10 多伦多大学医学系;加拿大安大略省多伦多 M5S 3H2 24 11 Unity Health;加拿大安大略省多伦多 M5B 1W8 25 12 西奈医疗系统;加拿大安大略省多伦多 M5G 1X5 26 13 大学健康网络;加拿大安大略省多伦多 M5G 2C4 27 14 多伦多大学分子遗传学系;加拿大安大略省多伦多 M5S 1A8 28 15 多伦多大学达拉拉纳公共卫生学院;加拿大安大略省多伦多 M5S 1A1 29 30 *通信地址:arinjay.banerjee@usask.ca (AB) 和 darryl.falzarano@usask.ca (DF)
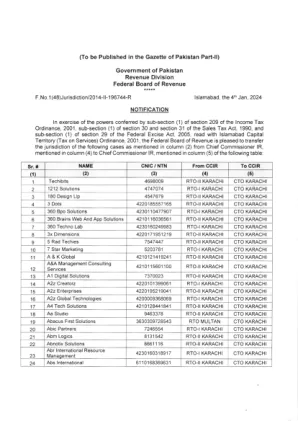
RTO II 卡拉奇 CTO 卡拉奇
CTO KARACHI 42301 34387986 Access Qore 26 CTO KARACHI RTO.II KARACHI 5084 889 Accounting Bookkeepers 27 CTO KARACHI RTO HYDERABAD 4130396508939 Accurasy (Smc Private) Limited CTO KARACHI RTO.I KARACHI 4230107616899 29 Accure Engineering CTO KARACH I RTO.II KARACHI A307300 Ace lnternational Marketing 30 4210115353445 Ace Marketing 31 9105620 Aciano Technologies 32 CTO KARACH I RTO-I KARACHI 681 1797 Adtechventure 33 CTO KARACHI MTO KARACHI 2241775 Technol les Advanced Research Proiects And Ltd 34 CTO KARACHI RTO-II KARACHI 4220112176555 Ag World CTO KARACH I RTO.II KARACHI 4220190426643 Agile Support Partners CTO KARACH I RTO-I KARACHI Ahmad And Qadeer Holdings CTO KARACHI RTO-II KARACHI 7505491 Aib Global 38 CTO KARACH I RTO-I KARACH I 3656120 Aim Viz CTO KARACHI RTO-II KARACHI 4320180634443 Aims International CTO KARACHI RTO-II KARACHI 4220149179843 Ak-Mark Solutions 41 CTO KARACH I RTO-II KARACHI 421 0182333893 Al Azam Group 42 CTO KARACHI RTO-II KARACHI Al Nahyan International 43 CTO KARACHI RTO-I KARACH I Alali Technologies 44 CTO KARACH I RTO-II KARACHI 4200004812925 Al-Ankaboot lnternational CTO KARACHI RTO-II KARACHI 7037629 Albor Tech 46 CTO KARACH I RTO-II KARACHI 4550103021629 Alfa Telemed 47 CTO KARACHI RTO-I KARACHI 4230119428317 Algoace CTO KARACHI RTO.I KARACH I 42301210347 53 49 CTO KARACHI 5220364565387 Ali Builders 50 CTO KARACHI RTO-II KARACH I 4250176525059 51 CTO KARACH I RTO-II KARACHI 4210107312205 Alp Bean CTO KARACHI RTO-II Karachi Alpha代表CTO KARACH I RTO-I KARACHI AL-REHMAN技术54 CTO KARACHI RTO IVIU LTAN 3240413959709 AM创意CTO COTO KARACHI KARACHI RTO-II RTO-II II KARACH I 4230113821381 CTO KARACH I RTO KARACH I RTO RACH IR 421110 TO-II KARACHI AMAXZA数字58 CTO KARACH I RTO-I KARACHI 4210154051793 AMBAS LNTERNATITAL 59 CTO KARACHI RTO-II KARACHI 7259030 AMCO AMCO AMCO LNTERMCO LNTERNTERM 60 60 COTO LNTERNTARTIC 35美国健康和供应 62 CTO 卡拉奇 RTO-I 卡拉奇 I 1a1'70'>7 Amfco lnternational bJ CTO 卡拉奇 RTO-II 卡拉奇 4220124224917 64 CTO 卡拉奇 4220136085876 Amotech Solutions

引用González-Iglesias E,Ochoa D,RománM,Soria-Chacartegui P,Martín-Vilchez S,Navares-GómezM
生物等效性临床试验涉及健康的志愿者,其血液检查必须在正常范围内,这对于胆红素和肝酶非常严格。吉尔伯特的综合征(GS)是与胆红素在肝脏中的代谢有关的良性遗传疾病(Düzenli等,2021)。胆红素是血红素分解代谢的最后产物,主要来自网状内皮系统中红细胞血红蛋白的崩溃(Memon等,2016)。胆红素消除是通过与葡萄糖酸结合将其转化为直接胆红素的(Gil andSąSiadek,2012)。由于GS患者的葡萄糖醛酸化水平降低,而未偶联的胆红素并非像共轭胆红素那样水溶性,因此不能将其排泄在胆汁中,患者患有未偶联的高胆汁纤维血症和轻度的高度脱节性高度,thoguluva chandrasekar et al al al al al al and and chandymirirubinia and。在健康的人中,胆红素的正常水平范围为0.1至1.2 mg/dl。但是,GS患者的水平通常为1.2至5.3 mg/dl(Gil andSąSiadek,2012年)。因此,由于怀疑任何肝病,胆红素水平升高的GS患者被排除在生物等效性研究之外,即使这种变化在临床上是微不足道的,并且众所周知,该综合征患者的肝酶没有改变(Moreno等人,1984; 1984; sidiib; sidorenko and teirenko and t.222222222) 2023)。这种变化称为等位基因UGT1A1 *28(RS3064744),以前被注释为RS34815109或RS34983651(Aronica等,2022)。gs患者在基因中具有变体,用于将未偶联的胆红素转化为共轭胆红素,尿苷二磷酸葡萄糖醛酸葡萄糖葡萄糖基转糖基转移酶1A1(Thogululuva chandrasekar等)。更具体地,它与该基因的启动子的短串联重复(Str)变化有关,该启动子包括将二核苷酸序列(TA)添加到转录启动序列A(TA)7 TAA中,将其转换为(TA)8 TAA(Horsfall等,2011; Thoguluva; Thoguluva Chandrasekar et a(Horsfall et al taa taa)。因此,具有这种变体使酶仅具有正常活性的30%。此外,当添加一个二核苷酸序列(A(TA)6 TAA)或UGT1A1*37时,当基因组中的该位置定义了其他等位基因,例如UGT1A1*36(a(ta)9 TAA)。UGT1A1*36的转录水平似乎高于UGT1A1*1,而UGT1A1*37似乎具有较低的水平(Gammal等,2016)。这些变体不太常见或可能取决于祖先的地理区域(Gammal等,2016)。并非每个具有等位基因UGT1A1 *28的人最终都会出现明显的症状,因为它取决于环境因素,例如身体压力,延长禁食,饮食不良,溶血反应,发热疾病和月经(Düzenli等,2021年)。UGT1A1 RS887829 C> t变体(UGT1A1 *80)因与UGT1A1 *28有可能的关系而进行了研究。已被描述为与UGT1A1*28的几乎完全连锁不平衡(R 2例如,在48小时内,降低热量摄取至400 kcal日记会增加胆红素浓度2至3倍。 GS通常出现在青春期早期,并且在男性中更频繁地诊断出,由于性类固醇浓度差异和雄性胆红素的产生较高而引起的女性(Thoguluva Chandrasekar等,2022年)。