XiaoMi-AI文件搜索系统
World File Search System35S

COVID-19疫苗对Delta变体的有效性(...
启动子。为了研究热诱导的indels是否是启动子特异性的,我们测试了三个常用的本构启动子35S,RPS5A和Zmubi以驱动Cas9表达[34-36]。我们筛选了每个启动子-CAS9组合的9-14独立分离T2种群,以两到四个不同的GRNA靶向PDS3。我们观察到3xHS处理后,PDS3表型的数量和/或严重程度增加了Zmubi-Cas9(6/9独立线),RPS5A-CAS9(14/14独立线),35S- Cas9(6/12独立线)和PCUBI-CAS9和PCUBI-CAS9(7/9 Independed Line Independent lineption;图S4)。尽管表型对于35S-CAS9线的表型相对温和,但我们的数据与其他报道相一致,即热应力会提高与驱动CAS9的启动子序列无关的ideel效率[17,18,26]。热休克提高了烟草中的基因组编辑效率

关于启动子在植物基因工程中使用的更新和观点
基因工程植物在农业中的应用多样,以增强食物和饲料的价值。基因工程旨在将具有理想特征的选定遗传区域引入空间和时间表达的目标植物中。启动子是负责通过识别RNA聚合酶调节转录因子(TF)来调节基因表达的关键要素。基于它们的识别和表达,RNA聚合酶分为RNA POL II和POL III启动子。 启动子活性和特定峰是调节转基因表达的两个主要参数。 由于使用构成启动子(例如Cauli-limpower Mosaic病毒(CAMV)35S)可能会导致对非目标生物或生态系统,可诱导/组织特异性启动子和/或RNA POL III启动子的不利影响,并为对控制的调节和最小值不良效应提供了多种机会。 除了它们在转基因表达中的作用外,还讨论了它们在合成生物学和基因组编辑中的影响。 本综述提供了有关迄今为止报道的启动子的优势和缺点的重要性,当前的前景和洞察力的最新信息,将有助于利用它们在努力中开发营养和农艺改善的转基因作物进行商业化。基于它们的识别和表达,RNA聚合酶分为RNA POL II和POL III启动子。启动子活性和特定峰是调节转基因表达的两个主要参数。由于使用构成启动子(例如Cauli-limpower Mosaic病毒(CAMV)35S)可能会导致对非目标生物或生态系统,可诱导/组织特异性启动子和/或RNA POL III启动子的不利影响,并为对控制的调节和最小值不良效应提供了多种机会。除了它们在转基因表达中的作用外,还讨论了它们在合成生物学和基因组编辑中的影响。本综述提供了有关迄今为止报道的启动子的优势和缺点的重要性,当前的前景和洞察力的最新信息,将有助于利用它们在努力中开发营养和农艺改善的转基因作物进行商业化。

在不牺牲特异性的情况下在生菜中建立有效的基因组编辑系统
CRISPR/Cas9 基因组编辑系统的效率在许多作物中仍然有限。利用强启动子来提高 Cas9 的表达水平是提高编辑效率的常用方法。然而,这些策略也增加了脱靶突变的风险。在这里,我们开发了一种新策略,利用内含子介导增强 (IME) 辅助的 35S 启动子来驱动 Cas9 和 sgRNA 在单个转录本中,通过适度增强 Cas9 和 sgRNA 的表达来提高编辑效率。此外,我们开发了另一种策略来富集高表达 Cas9 /sgRNA 的细胞,通过共表达发育调控基因 GRF5 ,这已被证明可以提高转化效率,并且来自这些细胞的转基因植物也表现出增强的编辑效率。该系统将莴苣(Lactuca sativa)中三个目标的基因组编辑效率从 14–28% 提高到 54–81%,且脱靶编辑效率没有增加。因此,我们建立了一种新的基因组编辑系统,该系统大大提高了目标编辑效率,且没有明显增加脱靶效应,可用于表征莴苣和其他作物中的目标基因。

使用BT Cry2a基因和印度品种Arka Vikas 的遗传转化使用BT Cry2a基因和印度品种Arka Vikas
南印度品种Arka Vikas的转基因番茄植物是使用农杆菌菌株EHA 105开发的,该菌株具有bt Cry2a基因,其中包含35S CAMV启动子,OCS终止剂和NPTII -NEPTI -NEPTI -NOPTII -abledable Marker,通过Agrobacterium Medimed -MediDied Transformation。进行了这项研究是为了改善南印度品种Arka Vikas的再生和转化方案。下胚基被用作由于较高的再生效率,通过PCR分子分析t 0生成中的推定转化体,用于t 0生成中的分子分析,并进行了定性ELISA方法,以用于BT蛋白表达,然后进行昆虫生物测定。昆虫生物测定研究,以筛选植物,并在后代进一步携带了用分子和表型特征表达良好耐药性的植物。实验结果得出的结论是,BT基因成功地部署在番茄品种中,并在实验室条件下对Helicoverpa Armigera的新生儿幼虫产生了抗性。这些结果表明转基因线在Helicoverpa Armigera的管理中有效地表达了大量的BT Cry2a蛋白。转基因T 1系的精确筛选对于获得单拷贝数植物非常重要,因为连续一代中BT蛋白的表达促进了将来该害虫的有效管理。

在番茄中应用CRISPR/CAS9介导的基因编辑
crispr-/cas9介导的基因编辑已在包括番茄在内的许多食品作物中证明。番茄(Solanum lycopersicum)既是重要的粮食作物,又是一种模型植物物种,已广泛用于研究基因功能,尤其是与水果生物学有关的植物。这种双重性在目的中与随时可用的资源(突变种群,基因组序列,转化方法)相结合,使番茄成为基因编辑的理想候选者。我们实验室通常使用的CRISPR/CAS9系统已应用于各种番茄基因型和野生物种solanum pimpinellium。矢量系统基于金门克隆技术。盒,该基因既赋予对卡纳米霉素的抗性,Kanamycin是由CAMV 35S启动子驱动的人类密码子驱动的Cas9,并在控制拟南芥U6 Polymerase促进剂的控制下引导RNA(GRNA)是组装成T-Dna的casss cass9。通常,我们设计了每个基因靶标的两个GRNA的CRISPR/CAS9构建体。但是,我们已经成功地包括多达八个grnas,以同时针对多个基因和区域。将CRISPR-/CAS9设计的构建体引入番茄中是通过基于基于NPTII基因的存在的培养基的培养基的培养基感染的转化方法来实现的。本章详细介绍了CRISPR/CAS9构建体和基因型分析(基于PCR的扩增子测序和T7核酸内切酶)的方法。
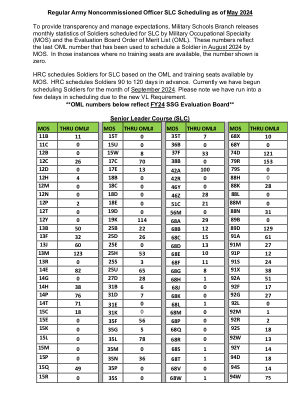
截至 2024 年 5 月的正规陆军士官 SLC 排程
MOS 穿过 OML# MOS 穿过 OML# MOS 穿过 OML# MOS 穿过 OML# 11B 11 15T 0 35T 7 68X 10 11C 0 15U 0 36B 0 68Y 0 12B 0 15W 8 37F 33 74D 121 12C 26 17C 70 38B 0 79R 153 12D 0 17E 13 42A 100 79S 0 12H 4 18B 0 42R 0 88H 0 12M 0 18C 0 46Y 0 88K 28 12N 0 18D 0 46Z 28 88L 0 12P 2 18E 0 51C 21 88M 0 12T 0 19D 0 56M 0 88N 31 12Y 0 19K 114 68A 29 89B 0 13B 50 25B 22 68B 12 89D 129 13F 32 25D 26 68C 15 91A 61 13J 60 25E 0 68D 13 91M 27 13M 123 25H 53 68E 10 91P 12 13R 0 25S 3 68F 11 91S 24 14E 82 25U 65 68G 8 91X 38 14G 0 27D 28 68H 1 92A 51 14H 38 31B 6 68J 0 92F 17 14P 76 31D 7 68K 0 92G 27 14T 71 31E 0 68L 1 92L 0 15C 18 31K 0 68M 0 92M 1 15E 0 35F 56 68P 0 92R 2 15K 0 35G 5 68Q 0 92S 18 15L 0 35L 78 68R 0 92W 13 15M 0 35M 0 68S 1 92Y 14 15P 0 35N 36 68T 1 94D 18 15Q 49 35P 0 68V 0 94S 14 15R 0 35S 0 68W 1 94W 75

94 个 CRISPR-Cas9 表达 T0 转基因烟草株系的基因编辑谱揭示了目标基因的高嵌合编辑频率
摘要:嵌合编辑是基因编辑中经常报道的技术。为了评估嵌合编辑的普遍情况,我们构建了携带标记β-葡萄糖醛酸酶基因(gusA)的转基因烟草品系,并将CRISPR-Cas9编辑载体引入转基因烟草品系中以敲除gusA,然后研究T0代及后续代中gusA的编辑效率。编辑载体携带一个由花椰菜花叶病毒35S启动子驱动的Cas9基因,以及两个引导RNA,gRNA1和gRNA2,分别由拟南芥U6(AtU6)和U3(AtU3)启动子驱动。两个gRNA被设计用于敲除gusA编码区的一个42个核苷酸的片段。利用农杆菌介导的转化和潮霉素选择将编辑载体转化到含有gusA的烟草叶中。使用抗潮霉素的独立 T 0 转基因株系通过组织化学 GUS 测定、聚合酶链式反应 (PCR) 和 PCR 扩增子的下一代测序来评估 gusA 编辑效率。94 个 T 0 转基因株系的靶序列图谱显示,这些株系是由未经编辑的细胞再生而来的,随后进行了编辑,并在再生期间或之后在这些株系中产生了嵌合编辑细胞。其中两个株系的 42 bp 核苷酸对的靶片段被去除。详细分析表明,在 4.3% 和 77.7% 的 T 0 转基因株系中分别发现了 AtU6-gRNA1 位点和 AtU3-gRNA2 位点的靶突变。为了解决 T 0 株系中编辑效率极低的问题,我们从嵌合株系进行了第二轮芽诱导,以提高获得所有或大多数细胞都经过编辑的株系的成功率。 T 0 转基因系中的突变谱为理解利用组成型表达的 CRISPR-Cas9 和 gRNA 进行植物细胞中的基因编辑提供了宝贵的信息。

开发模块化双生病毒载体以实现植物的高表达和基因靶向
* 通讯地址:电话:919-515-5729;电子邮件:jmalonso@ncsu.edu 摘要 病毒载体可以成为表达重组蛋白以及递送基因编辑机制的有用工具。尽管它们很有用,但这些工具的开发和后续优化通常是一个困难而繁琐的过程。因此,尽管已经做了大量工作来创建用于基因编辑和蛋白质表达的有用病毒载体,但对于如何最好地设计这些载体以用于特定应用,人们缺乏了解。例如,通常不清楚加入异源启动子序列或不同的病毒成分是否会改善货物表达或复制子积累。为了解决其中一些障碍,我们设计了一种基于双生病毒 - 甜菜卷叶病毒 (BCTV) 的 GoldenBraid (GB) 兼容病毒载体系统。该系统允许对各种报告构建体进行简单的模块化克隆。利用这种模块化克隆策略,我们比较了各种替代病毒载体架构。有趣的是,天然 BCTV 启动子的表现优于组成型 35S 启动子,而 BCTV 病毒体正义基因的去除则促进了报告基因的表达。有趣的是,这些修改对总复制子积累没有影响。这些结果表明了新的模块化基于 BCTV 的病毒载体在蛋白质表达和基因靶向应用方面的实用性,同时也揭示了可能为未来基于双生病毒的病毒载体架构提供信息的设计原则。我们预计,这种新模块化系统的推出将引发基于复制子的策略在植物蛋白质表达和基因编辑实验中的广泛应用。关键词:病毒载体、基因编辑、甜菜曲顶病毒、双生病毒、GoldenBraid、瞬时表达。简介病毒载体已被证明可用于各种生物技术应用,例如基因组编辑和蛋白质表达。基于双生病毒的病毒载体已用于递送基因组编辑酶,例如锌指核酸酶 (ZFN) 和 Cas9,以及用于同源定向修复 (HDR) 的修复模板 (RT) (Butler 等人,2016 年;Wang 等人,2017 年;Yu 等人,2020 年;Gil-Humanes 等人,2017 年;Dahan-Meir 等人,2018 年,Eini 等人,2022 年)。

蓖麻植物中的 CRISPR/Cas9 介导基因组编辑......
摘要 动机:CRISPR/Cas9 技术已被开发为最有效和最广泛使用的基因组编辑工具,用于修改众多植物的基因组,其中双链 DNA 中的 cas9 切割由单个向导 RNA(sgRNA)中包含的 20 个核苷酸序列驱动。然而,使用 CRISPR/Cas9 同时编辑多个目标仍然是该领域的技术挑战(Ma 等人,2014 年)。方法:在本研究中,使用 Golden Gate Assembly 克隆策略生成多个 CRISPR/cas9 编辑结构以用于蓖麻植物。模块化克隆系统使用 IIS 型酶在其识别位点外切割,从而允许有效组装具有兼容突出端的 DNA 片段,从而同时促进多个序列的正确取向(Engler 等人,2014 年)。我们的主要目标是获得一种遗传构建体,允许在同一个质粒载体中表达两个 sgRNA 和 cas9 核酸酶,以便通过农杆菌感染转化蓖麻。选择了两个针对 FAH12 蓖麻羟化酶的 CRISPR 靶标以避免可能的脱靶。这些靶标包含在 sgRNA 中并克隆到 0 级质粒中,每个质粒两侧都有 BsaI 酶的限制位点。Golden Gate 1 级反应包括几个 BsaI 消化和连接循环,将 U6 启动子与两个 sgRNA 分别组装到 1 级质粒中,两侧都有 BpiI 限制位点。同时,cas9 酶在双强 35S 启动子的控制下克隆,随后是来自 0 级质粒的胭脂碱合酶 (nosT) 终止子,包括这些元素,克隆到另一个 1 级质粒中,两侧也有 BpiI 限制位点。然后,用 BpiI 消化所有 1 级元件(U6-sgRNA1、U6-sgRNA2、2x35S-cas9-nosT)时,会出现兼容的突出端,这些突出端可以以正确的顺序和方向组装成 2 级结构。最终结果是 2 级质粒,其中包括 FAH12 羟化酶的 CRISPR/cas9 多重基因组编辑所需的所有元件。该构建体将转移到农杆菌中,以便以后进行蓖麻胚转化。