XiaoMi-AI文件搜索系统
World File Search SystemAAV

靶向整合素alpha v beta 6的工程AAV提出了跨物种的肌肉主义
使用自然衍生的AAV的当前与腺相关病毒(AAV)基因治疗受到非最佳组织靶向的限制。在治疗肌肉疾病(MD)时,通常需要高剂量,但会导致严重的不良反应。在这里,我们合理地设计了一种aav capsid,该capsid特异性地靶向绞肌以降低治疗剂量。我们将骨骼肌受体的人内整合素alphav beta6的结合基序计算整合到肝脏capsid中。与父母相比,设计的AAVS显示出更高的生产率和优质的肌肉转导。一种变体LICA1,示例可与其他肌肉AAV相比的肌肉转导,肝脏靶向降低。lica1在包括非人类灵长类动物在内的物种之间观察到了肌体性。因此,LICA1(而不是AAV9)有效地以低剂量(5E12 VG/kg)的两种小鼠MD模型(雄性小鼠)提供了治疗性转基因和改善的肌肉功能。这些结果强调了我们设计方法对MD基因疗法的AAV工程和LICA1变体的潜力。

cpg载体基因组对AAV基因治疗中CD8+ T细胞反应的影响
adeno相关的病毒(AAV)向量已成为体内基因替代疗法的首选平台,并代表了治疗单基因疾病(如血友病)的最有希望的策略之一。然而,对基因转移的免疫反应在临床试验中阻碍了人类基因治疗。在过去的十年中,很明显,先天免疫识别为诱导抗原特异性反应提供了信号,以针对载体或转基因产物产生。尤其是,TLR9识别对静脉细胞类树突状细胞(PDC)中载体的DNA基因组的识别已被鉴定为关键因素。来自临床试验和临床前研究的数据在矢量基因组中实施CpG基序,作为免疫反应的驱动因素,尤其是CD8 + T细胞激活的驱动因素。在这里,我们证明了AAV capsid特异性CD8 + T细胞的交叉化是否取决于XCR1 +

静脉注射AAV基因治疗用于治疗SOD1-ALS的静脉输送可提供NHP
超氧化物歧化酶1(SOD1)中的突变导致渐进性运动神经元通过有毒功能获得的特性丧失,并负责多达20%的家族性肌萎缩性侧面硬化症(ALS),或在美国所有ALS患者中大约2%的ALS患者评估了SOD1还原性降低的运动学策略,并且在所有ALS患者中均表现出了降低的运动,并改善了运动学,并改善了运动学的生存学,并具有SOD1患者的发展。表达突变体SOD1。最近对靶向SOD1的反义寡核苷酸的批准已进一步验证了SOD1作为治疗靶标。虽然减少SOD1的方法表现出不同程度的疗效,但它们依赖于无法实现最大治疗益处所必需的广泛,CNS范围的SOD1降低的直接CNS给药。我们先前报道了一系列体外和体内研究的结果,这些研究表明靶向SOD1的AAV基因治疗后,SOD1降低了。在G93A小鼠疾病模型中,我们在脊髓的整个尾声范围内证明了强大的SOD1敲低,运动性能的显着改善以及超出以前报道的核内核,肠内或肠内递送的生存延伸。在当前的研究中,我们将针对SOD1的高度有效的siRNA与静脉输送的,血脑屏障 - 透明剂tracer™capsid结合在一起,用于在NHP中进行评估。在2个月的生活期之后,我们观察到对脊髓和运动皮层的有利生物分布,从而显着降低了SOD1 mRNA。比静脉AAV递送中通常使用的新型衣壳固有的增强的BBB - 渗透率和自然的外围组织固有的固有的固定剂量,从而具有较低的剂量,从而产生了有利的安全性。这些结果表明,有效的SOD1 RNAi转基因与新型Tracer™衣壳的结合可显着减少临界脊髓和大脑区域中ALS中影响的SOD1 mRNA,并支持其持续的发展和发展到临床。

分泌的PD-L1通过局部和全身AAV基因治疗减轻小鼠的炎症性关节炎
结果:在表达SPD-L1(SHPD-L1)最佳亚型的AAV6载体的关节内注射后,与野生型PD-L1相比,观察到更大的效力,而关节炎改善所需的AAV6/SHPD-L1则较低。为了研究SPD-L1的全身表达的治疗作用,我们通过恢复轨道注射在CIA小鼠中施用AAV8/SHPD-L1基因治疗,并发现关节炎症和PAW肿胀的显着改善,在幼稚的小鼠中表现出相似的表型。在AAV8/ SHPD-L1处理的CIA小鼠中,总免疫球蛋白和抗胶原蛋白的特定抗体的水平低于对照组的抗体。在SHPD-L1处理的小鼠中,血液中促炎性细胞因子的水平也显着降低。此外,脾脏中的T细胞凋亡率在处理后的小鼠中增加了2倍。最后,我们通过肌肉注射研究了AAV/ SHPD-L1的治疗作用。注射AAV6/SHPD-L1后,pAW肿胀减少,关节炎症减少,血液中促炎细胞因子的降低和降低。SHPD-L1的治疗作用是通过用AAV载体肌肉处理来依赖于剂量的。

静脉注射AAV基因治疗用于治疗SOD1-ALS的静脉输送可提供NHP
在cynomolgus monkeys中静脉内给药Vy9323导致颈脊髓和腰椎脊髓中的大量载体基因组递送(A,B),并在颈椎和腰椎脊髓腹侧角组织(C,D)和Laser捕获的电动机(C,D)和Laser Captured captured的SOD1 mRNA减少。vy9323可大大降低颈脊髓中的SOD1 mRNA。黑色箭头代表表达许多SOD1 mRNA副本的细胞,红色箭头指示表达更少副本的细胞。在输送标记有效载荷和VY9323 CAPSID后,自动检测运动神经元转导,确定了85-94%的神经元在宫颈,胸腔和腰椎转导85-94%。

使用基于视紫红质的传感器
。cc-by-nc-nd 4.0国际许可证未通过同行评审获得证明)是作者/资助者,他已授予Biorxiv授予Biorxiv的许可,以永久显示预印本。它是此预印本的版权持有人(本版本发布于2024年5月5日。; https://doi.org/10.1101/2024.05.05.05.02.592211 doi:biorxiv Preprint

PBFT02 的非临床和早期临床开发,这是一种针对 GRN 突变 FTD 的 AAV 基因疗法(FTD- GRN)
## p < 0.01;### p < 0.005 vs WT;* p < 0.05,*** p < 0.005 vs Grn –/– + V,单因素方差分析,然后进行 Tukey 多重比较检验。缩写;GRN,颗粒蛋白基因;ICV,脑室内;V,载体;WT,野生型 Grn –/– 和 WT 小鼠(n=14-15/gp)ICV 给药 PBFT02 或载体(V)。基线对照是第 1 天未经治疗的小鼠。条形图:平均值 +/- SEM。
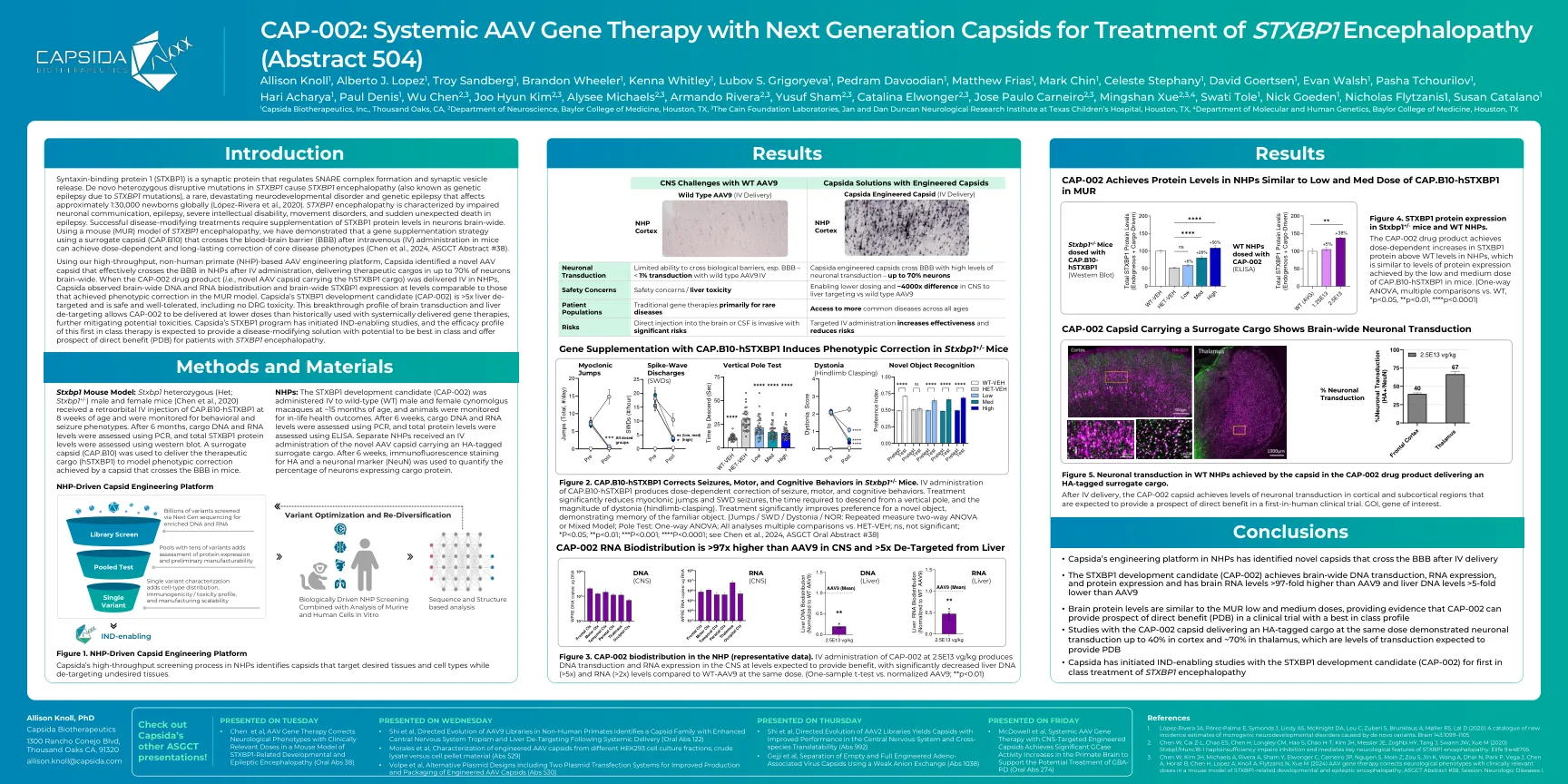
CAP-002:与下一代Capsids治疗STXBP1脑病的全身性AAV基因治疗(摘要504)
语法结合蛋白1(STXBP1)是一种突触蛋白,可调节SNARE复合物的形成和突触囊泡释放。STXBP1中的从头杂合性突变引起的STXBP1脑病(也称为STXBP1突变引起的遗传性癫痫),一种罕见的,破坏性的神经发育障碍和遗传性癫痫,影响全球范围1:30,000 Newborns Globally lul low low peepepepepepepepepez-ara eala and and newborns。STXBP1脑病的特征在于神经元交流,癫痫,严重的智力障碍,运动障碍和癫痫中突然意外死亡的特征。成功修改疾病的治疗需要在整个神经元中补充STXBP1蛋白水平。使用STXBP1脑病的小鼠(MUR)模型,我们证明,使用替代capsid(Cap.b10)的基因补充策略,该策略在小鼠中施用后跨液脑屏障(BBB)跨越血脑屏障(BBB),可以实现剂量依赖于剂量依赖于剂量的和长期的核心疾病(Ch)。 #38)。

使用 Cas9 和 AAV 研究全长 CFTR cDNA 替换引起的不利基因组和调控变化
一种替代全长 CFTR cDNA 的“通用策略”可治疗 99% 以上的囊性纤维化 (pwCF) 患者,无论他们的具体突变如何。基于 Cas9 的基因编辑被用于在气道基底干细胞的 CFTR 基因座处插入 CFTR cDNA 和截短的 CD19 (tCD19) 富集标签。该策略将 CFTR 功能恢复到非 CF 水平。在这里,我们通过评估 CFTR cDNA 插入后的基因组和调控变化来研究这种方法的安全性。首先通过使用 CAST-seq 量化基因重排来评估安全性。在验证编辑和富集的气道细胞中恢复的 CFTR 功能后,使用 ATAC-seq 表征 CFTR 基因座开放染色质谱。使用 scRNA-seq 评估编辑细胞中的再生潜力和差异基因表达。 CAST-seq 发现 0.01% 的等位基因发生易位,主要发生在非致癌脱靶位点,1% 的等位基因发生大量插入缺失。分化气道上皮细胞的开放染色质谱除 CFTR cDNA 和 tCD19 盒对应的区域外,没有出现明显变化,表明基因调控没有可检测到的变化。编辑后的干细胞产生的气道细胞类型与对照相同,基因表达的改变最小。总体而言,通用策略显示出轻微的不良基因组变化。

技术概述:使用SMRTBELL PREP套件3.0
Compatible with single - stranded AAV (ssAAV ) and self - complementary AAV (scAAV ) • AAV DNA sample extraction is performed using third - party methods• Multiplexing is performed using SMRTbell barcoded adapter plate 3.0 (102 - 009- 200)• A total of 1 μg of pooled AAV DNA is required for SMRTbell library preparation.每个单个AAV样本的输入DNA需求取决于多重水平,并且范围从