XiaoMi-AI文件搜索系统
World File Search SystemATF6

曲马多和/或氯胺酮在转移性去势抵抗性前列腺癌细胞系中重新用作潜在的抗癌药物
前列腺癌 (PCa) 进展为雄激素不依赖是死亡的主要原因。尽管所有转移性患者最初都对抗雄激素疗法有反应,但大多数患者在不到 2 年的时间内激素治疗失败。曲马多是一种阿片类激动剂,主要作用是治疗疼痛。氯胺酮是一种灵活的药物,具有广泛的临床用途。这项体外研究评估了曲马多和/或氯胺酮作为潜在抗癌药物的再利用。此外,还评估了这些药物对细胞死亡途径的影响。用曲马多和/或氯胺酮处理 PC-3 和 DU145 细胞系。通过实时 PCR 确定细胞凋亡、自噬、坏死性凋亡、部分死亡、内质网应激、Raf/MEK/ERK 通路和上皮-间质转化相关基因。目前的数据显示,与未经处理的癌细胞相比,经 TRA 和/或 KET 处理的 PC-3 细胞中大多数基因表达均上调,但 N-cadherin 除外,KET 对其下调幅度不大。另一方面,与未经处理的细胞相比,DU145 中所有处理过的细胞或未处理的癌细胞之间的基因表达差异不大,但 KET 显著上调了 ATG3、Beclin1 和 ATF6(分别为 P = 0.047、0.035 和 0.042),TRA 显著上调了 IRE1(P = 0.023),联合药物显著上调了 N-cadherin(P = 0.014)。与 DU145 细胞相比,PC-3 细胞对曲马多和/或氯胺酮的敏感性明显更高。ROS 诱导的细胞死亡途径可能是曲马多和氯胺酮对转移性 PCa 发挥抗癌作用的机制。针对细胞死亡途径是开发新型抗癌疗法的理想策略。

线粒体减少氨基氧霉素的成分1 p。 ...
缩写:165t,位于165位的苏氨酸(突变体); A165,位于165位的丙氨酸(野生型); AAV,腺相关病毒; ACTB,β-肌动蛋白; Alt,丙氨酸氨基转移酶; AST,天冬氨酸氨基转移酶; ATF6,激活转录因子6; CHX,环己酰亚胺; CQ,氯喹; DBEQ,Dibenzylquinazoline-2,4-二胺; ECL,增强的化学发光; ERAD,内质网相关降解; FACL4,脂肪酸-COA连接酶4; GCKR,葡萄糖酶调节剂; GWAS,全基因组协会研究; HMARC1,人线粒体减少的组件1; IP,免疫沉淀; IRE1,内切核酸酶肌醇提高酶1; ITR,反向终端重复;妈妈,线粒体相关的膜; MARC1,线粒体减少氨基氧霉素的成分1; MASLD,代谢功能障碍相关的脂肪分裂肝病; Mboat7,包含7的膜结合的O-酰基转移酶结构域; MMARC1,小鼠线粒体减少的成分1; ORO,油红色O染色; PERK,蛋白激酶R样性内质网(ER)激酶; PNPLA3,含patatin样磷脂酶结构域的蛋白3; RTA,相对总丰度; Ru,相对单位; SD,标准偏差; SDS,十二烷基硫酸钠; SDS-PAGE,十二烷基硫酸钠 - 聚丙烯酰胺凝胶电泳; SEM,平均值的标准误差; TM6SF2,跨膜6超家族成员2; UBC,泛素C; UBE2E1,泛素结合酶E2-E1; UBE3EC,泛素蛋白连接酶E3C; UPR,展开的蛋白质反应; UPS,泛素介导的蛋白酶体(降解)系统; VCP,含勇气的蛋白质。

SIRT1通过抑制内质网应激和NF-κB信号通路
沉默信息调节剂两个同源物1(SIRT1),一种NAD +依赖性组蛋白脱乙酰基酶,在无数生理过程中起关键的调节作用。越来越多的证据表明,SIRT1可以通过抑制内质网应力(ER)应力和核因子-κB(NF-κB)抗肿瘤信号信号传导途径来发挥代谢性疾病和神经退行性疾病的保护作用。本综述系统地阐明了SIRT1在调节ER应力和NF-κB途径中的分子机制和生物学意义。On one hand, SIRT1 can deacetylate key molecules in the ER stress pathway, such as glucose-regulated protein 78 (GRP78), X-box binding protein 1 (XBP1), PKR-like ER kinase (PERK), inositol- requiring enzyme 1 α (IRE1 α ), and activating transcription factor 6 (ATF6), thereby alleviating ER应力。另一方面,SIRT1可以直接或间接去除NF-κBp65亚基的乙酰化修饰,从而抑制其转录活性,从而衰减炎症反应。通过这些机制,SIRT1可以改善代谢疾病中的胰岛素抵抗,在缺血 - 再灌注损伤中发挥心脏保护作用,并减少神经退行性疾病中的神经元损害。然而,重要的是要注意,尽管这些发现是有希望的,但涉及的生物系统的复杂本质需要进一步研究,以完全揭示SIRT1的调节机制的复杂性。然而,了解SIRT1在ER应力和NF-κB途径上的调节机制非常重要,这对于扩大了我们对相关疾病发病机理的了解并探索针对SIRT1的新预防和治疗策略。
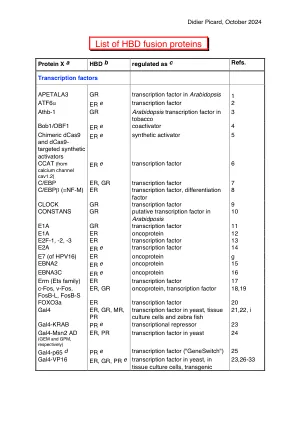
HBD 融合蛋白列表 - Picard 实验室 |
Picard, D. (2000)。通过与类固醇结合域融合实现蛋白质的翻译后调控。Methods Enzymol. 327 , 385-401。ER α HBD 融合在小鼠中的应用:Whitfield, J.、Littlewood, T.、Evan, GI 和 Soucek, L. (2015)。小鼠模型中的雌激素受体融合系统:可逆转换。Cold Spring Harb. Protoc. 2015 , 227-234。参考文献 1. Sablowski, RW 和 Meyerowitz, EM NO APICAL MERISTEM 的同源物是花同源基因 APETALA3/PISTILLATA 的直接靶标。Cell 92 , 93- 103 (1998)。 2. Thuerauf, DJ, Marcinko, M., Belmont, PJ 和 Glembotski, CC ATF6α 和 ATF6β 异构体特异性特征对内质网应激反应基因表达和细胞活力的影响。J. Biol. Chem. 282, 22865-22878 (2007)。3. Aoyama, T. 等人。拟南芥转录激活因子 Athb- 1 的异位表达改变了烟草叶细胞的命运。Plant Cell 7, 1773-1785 (1995)。4. Laumen, H., Nielsen, PJ 和 Wirth, T. BOB.1/OBF.1 辅激活因子对 B 细胞中八聚体依赖性转录至关重要。Eur. J. Immunol. 30, 458-469 (2000)。 5. Lu, J. 等人。用于转录激活和基因组编辑的多模式药物诱导 CRISPR/Cas9 装置。核酸研究。46,e25(2018)。6. Gomez-Ospina, N.、Tsuruta, F.、Barreto-Chang, O.、Hu, L. 和 Dolmetsch, R. L 型电压门控钙通道 ca(v)1.2 的 C 端编码转录因子。细胞 127,591-606(2006)。7. Umek, RM、Friedman, AD 和 McKnight, SL CCAAT 增强子结合蛋白:分化开关的组成部分。科学 251,288-292(1991)。 8. Müller, C., Kowenz-Leutz, E., Grieser-Ade, S., Graf, T. & Leutz, A. NF-M(鸡 C/EBP beta)诱导造血祖细胞系嗜酸性分化和凋亡。EMBO J. 14 , 6127-6135 (1995)。9. McDonald, MJ & Rosbash, M. 果蝇昼夜节律基因表达的微阵列分析和组织。Cell 107 , 567-578 (2001)。10. Simon, R., Igeno, MI & Coupland, G. 拟南芥花分生组织身份基因的激活。Nature 384 , 59-62 (1996)。 11. Picard, D., Salser, SJ & Yamamoto, KR 糖皮质激素受体类固醇结合域内可移动且可调节的失活功能。Cell 54 , 1073-1080 (1988)。12. Spitkovsky, D. 等人。腺病毒 E1A 在具有 E1A 依赖性条件性增殖的细胞系中对细胞周期蛋白基因表达的调节。J. Virol. 68 , 2206-2214 (1994)。13. Vigo, E. 等人。CDC25A 磷酸酶是 E2F 的靶标,是 E2F 有效诱导的 S 期所必需的。Mol. Cell. Biol. 19 , 6379-6395 (1999)。 14.Jones, ME, Kondo, M. & Zhuang, Y. A tamoxifen inducible knock-in allele for investigation of E2A function. BMC Dev. Biol. 9 , 51 (2009). 15.Zhao, B. et al. RNAs induced by Epstein-Barr virus Nuclear Antigen 2 in Lymphoblastoid cell lines. Proc. Natl. Acad. Sci. USA 103 , 1900-1905 (2006). 16. Maruo,S. 等人。Epstein–Barr 病毒核蛋白 EBNA3C 是淋巴母细胞细胞周期进程和生长维持所必需的。美国国家科学院院刊 103,19500-19505(2006 年)。