XiaoMi-AI文件搜索系统
World File Search SystemATPase
FHD-286,BRG1/BRM的有效抑制剂(...
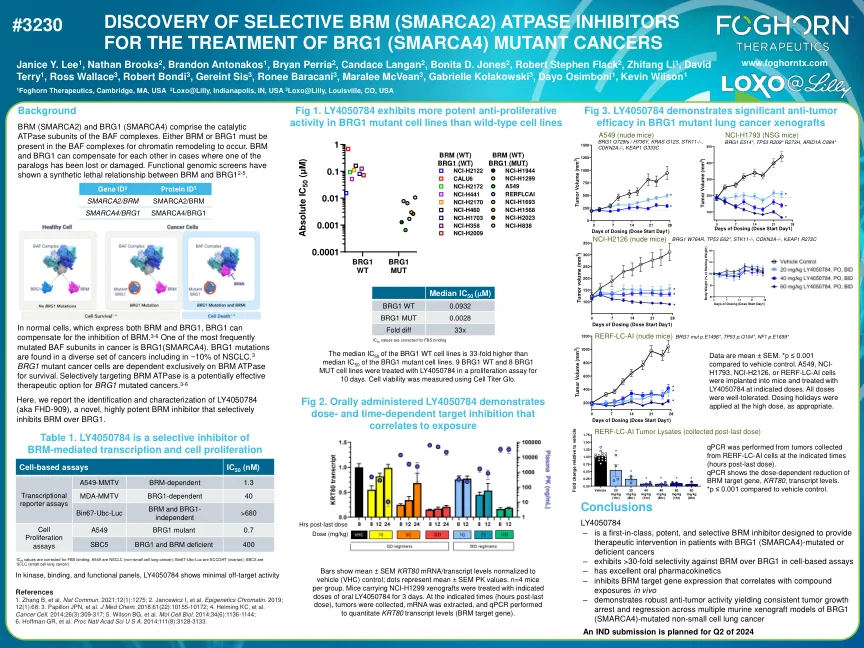
- 是一种旨在提供BRG1(SMARCA4)的患者(SMARCA4)的患者(SMARCA4)的治疗干预措施的一等,有效和选择性的BRM抑制剂 - 在基于细胞的测定中对BRG1的选择性> 30倍,在基于细胞的测定中表现出> 30倍的选择性 - 具有出色的口服药代动力学 - 抑制BRM的基因 - 抑制型号的export and comporiant in Comel and colly and colly and Incy Indect in Comel and Incy Indect in Comel and Incy Indect in Comel and Indecy Indect in Comel and Indect in Comel and Indectiv抗肿瘤活性在BRG1的多个鼠内异种移植模型(SMARCA4)中产生一致的肿瘤生长停滞和消退

通过抑制 ATPase RUVBL1/2 靶向胰腺癌中的 MYC 效应功能
摘要 目的 标志性致癌基因 MYC 驱动大多数肿瘤的进展,但小分子药物直接抑制 MYC 尚未进入临床试验。MYC 是一种依赖几种结合伙伴发挥作用的转录因子。因此,我们探索了通过胰腺导管腺癌 (PDAC) 中的相互作用组靶向 MYC 的可能性。 设计 为了在所有 MYC 结合伙伴中找出最合适的靶点,我们构建了一个靶向 shRNA 文库,并在培养的 PDAC 细胞和小鼠肿瘤中进行筛选。 结果 出乎意料的是,发现许多 MYC 结合伙伴对培养的 PDAC 细胞很重要,但在体内却不是必需的。然而,有些对自然环境中的肿瘤也是必不可少的,其中 ATPases RUVBL1 和 RUVBL2 排名第一。生长素-降解元系统降解 RUVBL1 导致培养的 PDAC 细胞停滞(而非未转化细胞),并导致小鼠的肿瘤完全消退,而此前免疫细胞浸润。从机制上讲,RUVBL1 是 MYC 建立致癌和免疫逃避基因表达所必需的,从而确定 RUVBL1/2 复合物是 MYC 驱动癌症中可用药的弱点。结论我们研究的一个含义是 PDAC 细胞依赖性受环境的强烈影响,因此应在体外和体内进行基因筛选。此外,生长素-降解元系统可应用于 PDAC 模型,从而允许在活体小鼠中进行靶标验证。最后,通过揭示 RUVBL1/2 复合物的核功能,我们的研究提出了一种使胰腺癌可能对免疫疗法敏感的药物策略。

通过抑制 ATPase RUVBL1/2 靶向胰腺癌中的 MYC 效应功能
摘要 目的 标志性致癌基因 MYC 驱动大多数肿瘤的进展,但小分子药物直接抑制 MYC 尚未进入临床试验。MYC 是一种依赖几种结合伙伴发挥作用的转录因子。因此,我们探索了通过胰腺导管腺癌 (PDAC) 中的相互作用组靶向 MYC 的可能性。 设计 为了在所有 MYC 结合伙伴中找出最合适的靶点,我们构建了一个靶向 shRNA 文库,并在培养的 PDAC 细胞和小鼠肿瘤中进行筛选。 结果 出乎意料的是,发现许多 MYC 结合伙伴对培养的 PDAC 细胞很重要,但在体内却不是必需的。然而,有些对自然环境中的肿瘤也是必不可少的,其中 ATPases RUVBL1 和 RUVBL2 排名第一。生长素-降解元系统降解 RUVBL1 导致培养的 PDAC 细胞停滞(而非未转化细胞),并导致小鼠的肿瘤完全消退,而此前免疫细胞浸润。从机制上讲,RUVBL1 是 MYC 建立致癌和免疫逃避基因表达所必需的,从而确定 RUVBL1/2 复合物是 MYC 驱动癌症中可用药的弱点。结论我们研究的一个含义是 PDAC 细胞依赖性受环境的强烈影响,因此应在体外和体内进行基因筛选。此外,生长素-降解元系统可应用于 PDAC 模型,从而允许在活体小鼠中进行靶标验证。最后,通过揭示 RUVBL1/2 复合物的核功能,我们的研究提出了一种使胰腺癌可能对免疫疗法敏感的药物策略。

新型 ATPase 抑制剂家族对具有高 NAT2 活性的结肠直肠癌细胞具有增强的细胞毒性
杂合性缺失 (LOH) 是癌症基因组的一个标志性特征,可减少等位基因变异,从而产生可用于治疗目的的肿瘤特异性弱点。我们之前报告过,8p22 处的 LOH 导致药物代谢芳胺 N -乙酰转移酶 2 (NAT2) 活性丧失,可作为结直肠癌 (CRC) 的附带致死抗癌疗法的靶点。在这里,我们报告了一种新型化合物 CBK034026C,它对具有高 NAT2 活性的 CRC 细胞表现出特异性毒性。连接图分析显示,CBK034026C 引发了与 ATPase 抑制剂相关的反应模式。与 Na + /K + -ATPase 的强效抑制剂乌巴因类似,CBK034026C 激活了 Nf-kB 通路。进一步的代谢组学分析表明,在具有高 NAT2 活性的 CRC 细胞中,与抗氧化防御和线粒体代谢相关的通路下调,从而削弱了对 CBK034026C 诱导的氧化应激的保护性反应。鉴定出一种针对由 NAT2 活性引起的代谢脆弱性的小分子,为开发抗癌药物提供了新途径。

口服生物可利用的 mSWI/SNF ATPase 降解剂的开发和前列腺癌的获得性耐药机制
2 美国密歇根州安娜堡密歇根大学病理学系 3 中国湖南省长沙市中南大学湘雅医院泌尿外科 4 美国密歇根州安娜堡密歇根大学医学科学家培训项目 5 美国密歇根州安娜堡密歇根大学细胞与分子生物学项目 6 美国密歇根州安娜堡密歇根大学罗格尔癌症中心 7 美国密歇根州安娜堡密歇根大学公共卫生学院生物统计学系 8 美国密歇根州安娜堡密歇根大学霍华德休斯医学研究所 9 印度班加罗尔 Aurigene Oncology Limited 10 美国密歇根州安娜堡密歇根大学分子与整合生理学系 11 美国密歇根州安娜堡密歇根大学内科系、胃肠病学分部

分析城市能源以实现净零能源社区
在转基因作物中表达的外源蛋白的细胞定位不仅决定了其稳定性,而且还决定了它们对作物生长和发育的影响,包括在压力条件下;然而,潜在的分子机制仍然未知。在这里,我们通过亚细胞定位,免疫组织化学,免疫流畅和蛋白质印迹分析确定了抗昆虫的转基因水稻huahui-1(HH1)细胞中外源表达的Cry1Ab/c蛋白的细胞分布。通过酵母两杂交,双分子分子荧光互补(BIFC)和辅助药物分析研究了CRY1AB/C蛋白与初筛选的内源性质膜Ca 2+ ATPase之间的相互作用。通过比较CRY1AB/C和Ca 2+ ATPase之间的细胞定位和相互作用位点分析了潜在的相互作用机制。表型指数和Ca 2+ -ATPase活性在转基因HH1和父母线Minghui-63在无压力和盐压力的条件下确定,可以由CRY1AB/C-Ca 2+ -ATPase相互作用调节。结果表明,Cry1ab/C不仅分布在细胞质和核中,而且还分布在质膜上,在质膜上与质膜Ca 2+ -ATPase相互作用。通过BIFC实验,这种相互作用部分保留了细胞核中的质膜蛋白Ca 2+ ATPase,因此可能会通过改变蛋白质的细胞位置来影响膜上Ca 2+ -ATPase活性。一致地,我们的结果证实了转基因HH1中Cry1ab/c的存在导致Ca 2+ -ATPase活性的降低,并对植物表型造成不利影响,包括显着降低的植物高度和生物量,与亲属MH63相比;并且这些有害作用在盐应力条件下更明显,从而影响转基因

发现选择性BRM(SMARCA2)ATPase抑制剂...
注意: *EP300损失可以通过以下任何遗传改变来弥补:深层缺失,杂合性丧失,高影响力的高影响突变,多重影响突变,一个中等影响突变和一个或多个中等或高影响突变,中度或高影响突变,中等影响突变,伴有杂合性丧失,杂合性损失或低基因表达。来源:DRG流行病学数据,TCGA分析

Bafilomycins:一类来自微生物、动物细胞和植物细胞的膜 ATPase 抑制剂
在原核生物和真核生物中,大多数已鉴定的离子泵 ATPase 属于以下三种结构类型之一。(i)F1Fo ATPase(F 型)存在于线粒体内膜(2)、叶绿体类囊体膜(3)和细菌细胞质膜(4)中。(ii)E1E2 ATPase(P 型)存在于真菌(5)、植物(6)和动物的细胞质膜中[包括 Na',K4-ATPase(7)和 H +,K + -ATPase(8)],以及肌细胞的肌浆网(Ca 2+-ATPase)(9)和细菌细胞质膜(K+-ATPase)(10,11)。 (iii) 已鉴定出第三类 ATPase(V 型),并从真菌和植物液泡(参考文献 12 及其中的参考文献)、包被囊泡(13、14)和嗜铬颗粒(15、16)的膜中部分纯化。正如 Mellman 等人(17)所建议的,我们使用术语“液泡 ATPase”来指代第三类 ATPase。F1Fo ATPase 通常使用 H+ 的电化学梯度(18)或偶尔使用 Na+ 梯度(19)来合成 ATP。这种类型的酶也表现出 ATPase 活性,在某些情况下仅在用蛋白酶活化后才表现出 ATPase 活性(20)。叠氮化物和 N,N'-二环己基碳二酰亚胺可抑制 F1Fo ATPase 的酶活性;寡霉素也可抑制线粒体 ATPase(21)。在 E1E2 ATPases 中,ATP 水解释放的能量与阳离子跨膜转运偶联。酶循环通过构象状态,包括形成磷酸化中间体。酶活性不受叠氮化物或寡霉素的影响,但被钒酸盐特异性抑制,在大多数情况下被 N-乙基马来酰亚胺和异硫氰酸荧光素抑制,而对于 Na4 ,K4-ATPase,则被乌巴因抑制 (5-11)。液泡 ATPases 似乎会水解 ATP,产生质子梯度,用于酸化细胞内区室 (12、17、22)。这组 ATP 酶因其抑制剂特异性而与其他两组 ATP 酶区分开来。液泡 ATPase 不受叠氮化物、寡霉素、钒酸盐或乌巴因的抑制。相反,

肌球蛋白和其他能量转换 ATP 酶
摘要:本文旨在记录超分子复合物(如马达、泵和时钟 ATPase)中的能量转换和调节相互作用。使用自旋标记电子顺磁共振 (EPR) 光谱通过运动和距离测量来表征动力学和结构特征。特别是,我们重点研究了肌球蛋白 ATPase 与肌动蛋白-肌钙蛋白-原肌球蛋白、神经驱动蛋白 ATPase 与微管、P 型离子动力 ATPase 和蓝藻时钟 ATPase。最后,我们描述了各种能量转换系统的分子机制之间的关系或共同原理,以及柔性元件从一种状态到另一种状态的大规模热结构转变如何先于随后的不可逆化学反应。

经验能量图谱揭示蛋白酶体在多肽转运中的机制
摘要 AAA+ 家族中的环状 ATPase 复合物执行多种细胞功能,这些功能需要其各个 ATPase 亚基的构象转变之间的协调(Erzberger 和 Berger,2006 年;Puchades 等人,2020 年)。如何通过这些协调运动捕获 ATP 水解产生的能量来做机械功尚不清楚。在本研究中,我们开发了一种新方法,用于基于互补的结构和动力学测量来描绘蛋白酶体异六聚体 ATPase 复合物的核苷酸依赖性自由能景观 (FEL)。我们使用 FEL 模拟蛋白酶体的动力学并定量评估预测的结构和动力学特性。FEL 模型预测与本研究和以前研究中的广泛实验观察结果一致,并提出了蛋白酶体 ATPase 的新型机制特征。我们发现 ATPase 亚基的协同运动源自 ATPase 六聚体的设计,该设计要求每个核苷酸结合状态具有独特的最小自由能。ATP 水解通过触发 ATPase 复合物的能量耗散构象转变来决定底物转位的方向。