XiaoMi-AI文件搜索系统
World File Search SystemATPase

与ATP1A3相关的遗传改变的动物模型...
在过去20年中,特别是随着外显子组测序技术的出现,常染色体显性和从头突变,编码Na +,K + -ATPase(NKAα3)泵的Na +,K + -ATPase(NKAα3)泵的神经元特异性α3亚基,ATP1A3,ATP1A3,ATP1A3,已被确定为photirogical conterypic nequim nequarum os of neuromy os of -nequim os of nequim of nequim os ne nequim,ATP1A3的这些等位基因疾病包括(在严重性/残疾和儿童发育中发作的近似顺序):Polymicrogyria;童年的交替偏瘫;小脑共济失调,蛋白酶,PES洞穴,视神经萎缩和感觉神经性听力损失综合征;用小脑共济失调复发脑病;和快速发作的肌张力障碍 - 帕金森主义。一些患者呈现中间,非典型或组合表型。由于这些疾病目前很难治疗,因此对更有效的疗法的需求未满足。分子机制通过ATP1A3中突变导致广泛的神经系统症状的分子机制知之甚少。然而,使用遗传改变模型生物的体内比较研究可以洞悉NKAα3中引起疾病突变的生物学后果。在此,我们回顾了用于研究ATP1A3相关疾病的现有小鼠,斑马鱼,果蝇和Caenorhabditis模型,并讨论了他们对了解疾病机制和新型治疗学发展的潜在贡献。

直接和延迟的超敏反应...
Proton pump inhibitors (PPIs) are widely pre scribed for managing acid-related gastrointestinal con ditions such as gastric and duodenal ulcers, dyspepsia, gastroesophageal reflux disease (GERD), Zollinger-El lison syndrome, Helicobacter pylori ( H. pylori ) erad ication, and the prevention and management of与非甾体类抗炎药(NSAIDS)相关的溃疡(1)。这些药物通过不可逆地抑制胃顶细胞中的H⁺/k⁺-ATPase酶来抑制胃酸促进(2)。尽管通常被认为是安全有效的,但PPI与一系列不良反应相关联(1)。其中,超敏反应引起了重大的临床关注,其中包括从轻度皮肤病症状到严重的全身并发症的一系列表现(3)。

引用纸浆版本(APA):Cleary,S。R.,Seflova,J.,Cho,E.E.,E.E.,Bisht,K.,Khandelia,H.,Espinoza-Fonseca,L.M。,L. M.,&Robia,S.L。(2024)。
磷兰班(PLB)是一种跨膜小肽,可调节心脏肌肉中的肌质网Ca 2+ -ATPase(SERCA),但这种调节的物理机制仍然很熟悉。PLB降低了活性SERCA的Ca 2+敏感性,从而增加了泵循环所需的Ca 2+浓度。然而,当不存在ATP时,PLB不会降低Ca 2+与SERCA的结合,这表明PLB不会抑制SERCA Ca 2+ afintient。对这些看似冲突的结果的主要解释是,PLB在与Ca 2+结合相关的SERCA酶促循环中的转变减慢了转运Ca 2+的依赖性,而不会实际影响Ca 2+协调位点的等电数。在这里,我们考虑了另一个假设,即在没有ATP的情况下,Ca 2+结合的测量可忽略核苷酸结合的重要变构效应,从而增加了SERCA Ca 2+结合效果。我们推测PLB通过逆转这种同义来抑制SERCA。为了测试这一点,我们使用了荧光的SERCA生物传感器来量化非循环SERCA的Ca 2+在存在和不存在不可用的ATP-ANALOG AMPPCP的情况下。核苷酸激活增加了SERCA Ca 2+的原性,并且通过PLB的共表达逆转了这种效果。有趣的是,在没有核苷酸的情况下,PLB对Ca 2+的原性没有影响。这些结果调解了先前的ATPase分析与Ca 2+结合测定的冲突观察结果。此外,SERCA的结构分析揭示了连接ATP和Ca 2+结合位点的新型变构途径。我们提出的这一途径被PLB结合所破坏。因此,PLB通过通过ATP中断泵的变构激活而降低了SERCA的平衡Ca 2+。因此,PLB通过通过ATP中断泵的变构激活而降低了SERCA的平衡Ca 2+。

质子泵:质子抽水的分子机制,抑制剂和激活因子
蛋白质分子机器,也称为质子泵,是生物膜中最重要的元素。这些是膜蛋白,在所有生物体(包括某些病毒)中广泛代表和分布。他们有能力通过将质子从膜的一侧转移到另一侧来创建和维持电化学质子梯度。质子泵分为各种大型类别,它们在不同的能源的使用方面有所不同,每个能源具有不同的多肽组成和进化起源。蛋白质泵中泵送质子的自由能的来源可能是:富含能量的代谢物的化学能(F.E.,质子ATPases中),来自具有较低氧化还原电位的化合物的电子转移能量(在线粒体呼吸链链中)和光能(F.E.,f.e.,f.e.,f.e.,在视野蛋白质中)。质子泵中质子的转移通常是电源的。然而,也有同样重要的,甚至可能更重要的非电原质质子泵,例如胃粘膜的氢 - 氯荷ATPase或H + /K + ATPase,这主要负责胃含量的酸性胃含量。题为“质子泵:质子泵的抑制剂和激活因子”的新特刊,总共包括六项贡献:四个原始文章和2个评论。Siletsky S.A.和Borisov V.B.的评论[1]分析了末端呼吸氧化酶的活性位点中氧中间体的最新结构和功能研究,催化循环的特征以及这些Engymes的活性位点的特性。这些文章和评论提供了与质子泵有关的新信息,首先要了解它们催化的反应机制的基础知识,它们在细胞生理学方面的重要性以及细胞内信号传导的分子机制,并以其在医学中的应用而结束。尽管贡献不足,但它们仍涉及广泛的基本问题和应用问题,并提供了新信息:有关特定蛋白质质子泵的分子机制和催化特征(尤其是细胞色素氧化酶和ATP合成酶);关于细胞生理学的特征以及涉及质子泵的信号转导的调节和机制;以及关于使用药物的分子医学研究 - 胃H + /K + ATPase的质子泵的抑制剂。末端呼吸氧化酶在功能上相似但在结构和进化上包括两个主要不同的超家族:血红素 - 波波氧化酶(HCOS,包括线粒体的细胞色素氧化酶(COX))和BD -type type type cytotromes。所有这些都通过将氧气还原为水的四电子还原的催化反应结合在一起,该反应在没有活性位点的潜在危险活性活性氧(ROS)的形成和释放的情况下进行。这些真核生物和原核生物的这些膜酶转化了电子从细胞色素或奎尼尔转移到分子氧向跨膜质子梯度转移的化学键的能量。迄今为止,具有原子分辨率的三维结构与BD型氧化酶相反,HCOS不仅通过从膜的不同侧转移到催化中心,而且还因为氧化还原偶联的定向质子通过膜泵送的独特能力而产生质子动力。

内耳发育中依赖于ATP的染色质重塑
摘要:在转录,DNA复制和修复过程中,染色质结构经常进行调节以揭示特定的遗传区域并允许进入DNA相互作用的酶。ATP依赖性染色质重塑络合物使用ATP水解的能量通过重新定位和重新排列核小体来修饰染色质结构。这些复合物由保守的SNF2(催化ATPase亚基)定义,并分为四个家庭:CHD,SWI/SNF,ISWI和INO80。ATP依赖性染色质重塑者对于调节包括内耳在内的许多器官的发育和干细胞生物学至关重要。此外,编码为染色质重塑者一部分的蛋白质的基因突变已与许多神经感觉聋的情况有关。在这篇综述中,我们描述了这些复合物的组成,结构和功能活性,并讨论它们如何促进听力和神经感觉的耳聋。


剖析成年神经干细胞的时空多样性突触囊泡的分子结构-MDC存储库
在这项研究中,Kravčenko及其同事提高了我们对突触囊泡(SVS)(SVS)的理解,这对于神经递质的存储和释放至关重要。采用冷冻电子断层扫描,该研究表征了SV蛋白的多样性,其中包括SV表面上的小蛋白,内部的细长蛋白,以及随机分布在SVS表面的大V -ATP酶。v - ATPase结构显示出另一种跨膜相互作用伴侣突触素。这项研究在网格蛋白涂层的网状蛋白笼中发现了v- ATPases,并在囊泡上部分组装了网状蛋白涂层,并在神经元内和神经元内部,提供了对其结构对称性的见解。此外,该研究确定了细胞膜附近没有囊泡的网状蛋白篮。这些发现突出了SV的复杂分子结构,提供了广泛的透视图并补充了传统的蛋白质组学分析和荧光显微镜。

thapsigargin - 从传统医学到抗癌药
摘要:倍半萜烯内酯Thapsigargin是一种植物化学物质,在Thapsia L.的地中海植物的根和成果中发现,这些物种已在民间医学中用于治疗风湿性疼痛,肺部疾病,肺部疾病和女性不孕症。最近发现Thapsigargin是一种有效的细胞毒素,可通过抑制肌胞浆/内质网状Ca 2+ ATPase(SERCA)泵来诱导凋亡,这对于细胞生存能力是必需的。这种生物学活性鼓励对使用Thapsigargin作为一种新型抗肿瘤剂的研究,但是由于该化合物对正常细胞的毒性较高而受到阻碍。在这篇综述中,我们总结了有关thapsigargin作用的生物学活性和分子机制的最新知识,以及在合成不太毒性的thapsigargin衍生物中的进步,这些衍生物被开发为新型抗癌药物。
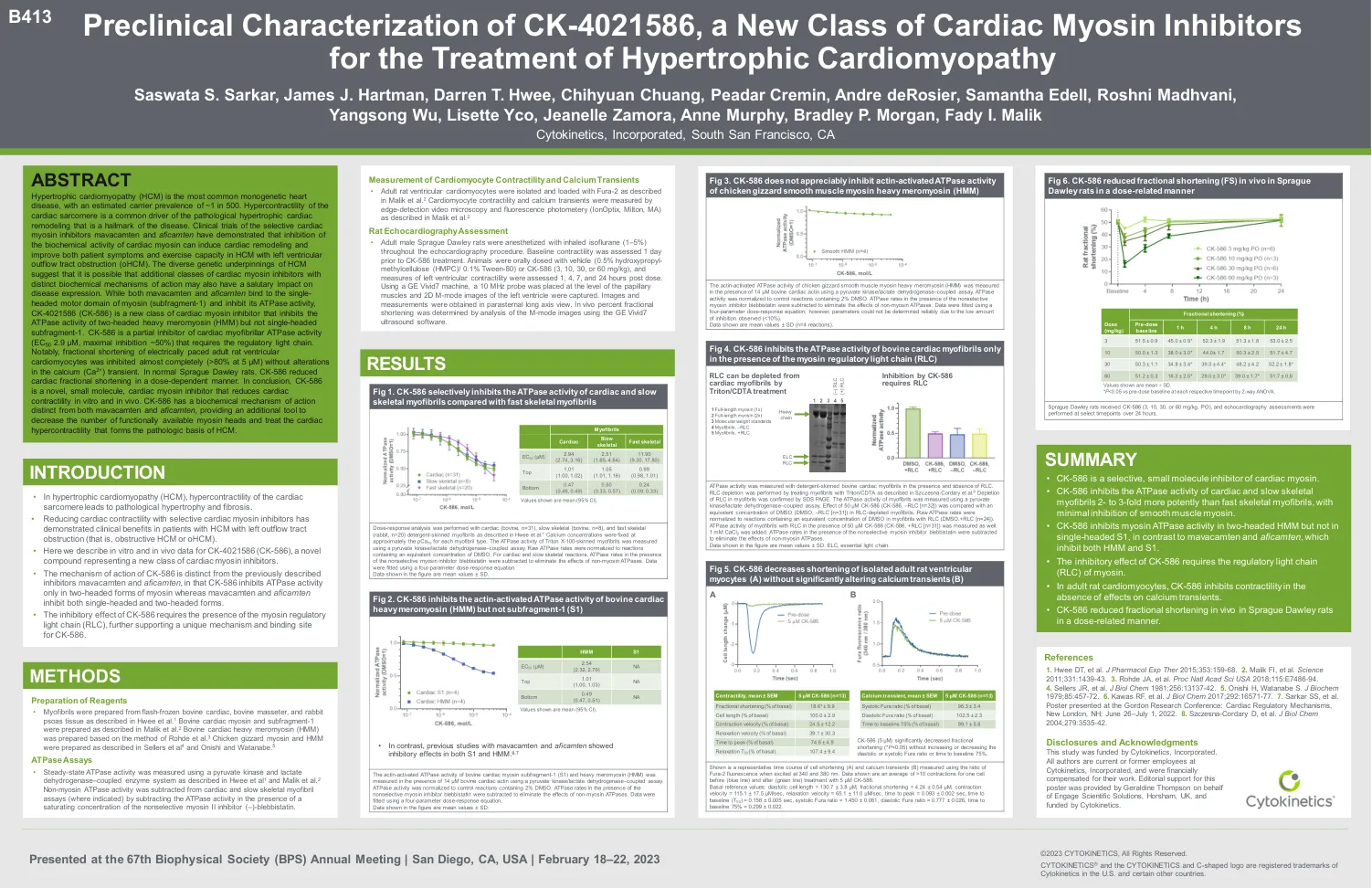
CK-4021586的临床前表征,一种新的心脏肌球蛋白抑制剂,用于治疗肥厚性心肌病
•在肥厚性心肌病(HCM)中,心脏肉瘤的超收缩性导致病理肥大和纤维化。•使用选择性心脏肌球蛋白抑制剂来降低心脏收缩性,显示出患有左流道阻塞的HCM患者的临床益处(即阻塞性HCM或OHCM)。•在这里,我们描述了CK-4021586(CK-586)的体外和体内数据,这是一种新型化合物,代表了一种新的心脏肌球蛋白抑制剂。•CK-586的作用机理与先前描述的抑制剂Mavacamten和Aficamten不同,因为CK-586仅以两种肌球蛋白的形式抑制ATPase活性,而Mavacamten和Aficamten和Aficamten均抑制了单头和两头的形式。•CK-586的抑制作用需要肌球蛋白调节轻链(RLC),进一步支持CK-586的独特机制和结合位点。

治疗万古霉素耐药肠球菌 (VRE) 感染的途径
研究小组假设,Na+转运VoV1-ATPase可能是VRE存活的重要酶。这种酶起到钠泵的作用,在肠道的碱性环境中将Na+输出出细胞,从而维持体内平衡并使细菌生长(图1a)。这种蛋白质存在于多种能在碱性环境下生长的致病菌中,但在动物、植物以及乳酸杆菌、双歧杆菌等有益菌中却不存在,因此抑制该蛋白质的化合物有望成为新型抗菌药物。 因此,我们假设,如果我们能够找到一种化合物来抑制这种 Na + 转运 V o V 1 -ATPase 的功能,我们也许能够抑制 VRE 的增殖,并且我们从广泛的化合物库中寻找抑制剂。