XiaoMi-AI文件搜索系统
World File Search SystemAda

美国残疾人法案 (ADA) 人行道过渡...
简介 2 人行道设计规范、标准和实践 2 公共工程路权范围内的街道和人行道改进 4 街道路权范围内现有人行道的清单 6 每种结构类型需要收集的清单信息 7 解决 ADA 合规性的改进计划 8 负责计划实施的官员 10 申诉程序 10 公众参与和过渡计划修订 12 附录 A. 城市街道、雨水、排水和污水管道人行道标准详情 B. MoDOT ADA 检查表 C. 城市完整街道和完整交叉口政策 D. 已完成的路缘坡道改进 E. 已完成的道路改进项目 F. 已完成的人行道改进项目 G. 已完成的信号人行横道改进项目 H. 已完成的公交候车亭安装项目 I. 清单和优先顺序图 J. 清单清单和排名 K. 清单摘要 L. 拟议的更正建议在城市财产上建造的结构,信号人行横道和公交候车亭 M. 拟议的包含行人设施改进的 CIP 项目 N. 与沥青覆盖层项目相关的拟议路缘坡道改进 O. 拟议的公交候车亭安装项目 P. ADA 歧视投诉表

2025 年共同立场:ADA 战略计划
75% 的选民在每个能力领域的表现至少为合格(5 分制中的 3 分)。 75% 的选民在所有能力领域的平均表现超过合格(5 分制中的 4 分)。 ADA 每年将完成 80-90% 的运营计划。

美国残疾人法案 ADA 过渡计划
第 1 节 - 一般信息 ................................................................................................................................................ 1 第 2 节 - 公共通行权 .............................................................................................................................................. 3 第 3 节 - 建筑和相关场地要素 ............................................................................................................................ 8 第 4 节 - 通讯 ...................................................................................................................................................... 9 第 5 节 - 交通 ...................................................................................................................................................... 10
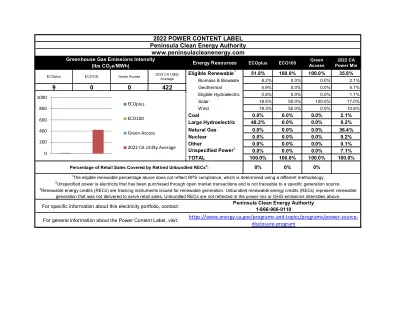
06 LOCA2混合降级 - 加利福尼亚能源委员会2022 PCL半岛清洁能源管理局ADA
1上述合格的可再生级别不反映RPS的依从性,这是使用不同方法确定的。2未指定的电力是通过公开市场交易购买的电力,无法追溯到特定的一代来源。3可再生能源信用(REC)正在跟踪为可再生生成发行的工具。捆绑可再生能源信用(REC)代表可再生能源

_AMERICANS残疾人法(ADA)标题2投诉过程
步骤4:密歇根州民权部(MDCR)审查。如果您和MSP ADA协调员无法同意决议,协调员将发送书面摘要,通常包括:(1)您的投诉说明; (2)提出的任何决议的摘要; (3)解决无法解决的问题的声明。如果您的投诉未解决,则可以要求MDCR进行进一步审查。您可以在收到协调员的非分辨率通知后的10个工作日内向MDCR提出任何请求,通过将您的原始投诉的副本和MSP摘要发送到mdcrservicecenter@mi.gov。
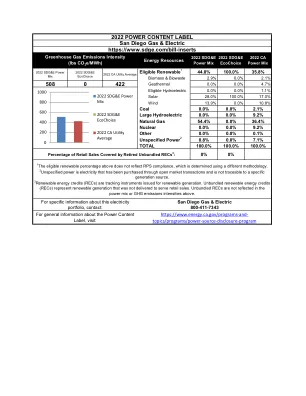
2022 PCL圣地亚哥天然气和电动ADA
Wind 13.9% 0.0% 10.8% Coal 0.0% 0.0% 2.1% Large Hydroelectric 0.0% 0.0% 9.2% Natural Gas 54.4% 0.0% 36.4% Nuclear 0.0% 0.0% 9.2% Other 0.0% 0.0% 0.1% Unspecified Power 2 0.8% 0.0% 7.1% TOTAL 100.0% 100.0% 100.0%

第76大道SE中间越过和ADA改进...
步行,骑自行车和滚动是默瑟岛(Mercer Island)的常见旅行模式,尤其是在市中心。随着今年晚些时候East Link Light火车站的开放,预计将会有更多非机动交通。第76大街SE中间街区和ADA改进项目的重点是升级现有设施,以便为市中心的所有居民和访客提供访问现有设施。该市的ADA过渡计划在2022年被市议会通过。本计划有助于指导未来的计划和在整个城市进行必要的可访问性改进。在2023 - 2024年,由ADA过渡计划完成的第一个项目完成了,在市中心建造了11个符合ADA的坡道。继续进入2025 - 2026年及以后,员工正在追求更多的ADA改进项目。位于市中心的心脏地带,现有的中间街区在7620 SE 27街和2690 76 Th Avenue SE之间的中心交叉处在给定的工作日中为近300名行人提供服务。交叉路口的相邻目的地包括零售,住房和过境设施。计划在此常用的中间越野上安装一个新的RRFB,以提高行人的能见度。第76大街SE中期交叉和ADA改进项目结合了该市六年的运输改进计划中的两个项目:76 Th Avenue SE SE中部街区穿越改进和ADA过渡计划实施。项目总预算为$ 850,368。工作范围

2024 年 ADA 护理标准:有哪些新内容?
美国糖尿病协会 (ADA) 的糖尿病护理标准是“现行指南”,随着重大新证据的出现,该标准全年都会更新,每年 1 月都会发布完整更新。本概览情况说明书重点介绍了 2024 年 1 月发布的关键变化。尽管标准的某些部分反映了美国的实践,但糖尿病护理正在全球范围内变得更加统一,因此大多数建议都与英国的医疗保健专业人员相关。这里重点介绍的变化主要针对与初级和社区护理团队相关的变化。

亨茨维尔地区 MPO ADA 过渡计划......
第六条声明 亨茨维尔地区大都会规划组织 (MPO) 致力于遵守 1964 年《民权法案》第六条、1987 年《民权恢复法案》以及所有相关规则和法规。亨茨维尔地区 MPO 保证,任何个人或群体都不会因种族、肤色、年龄、残疾、国籍、性别或收入状况而被排除在亨茨维尔地区 MPO 管理的所有计划、服务或活动之外,或被剥夺其福利,或受到其他歧视,无论这些计划和活动是否由联邦政府资助。亨茨维尔地区 MPO 的政策还包括确保其所有计划、政策和其他活动不会对少数族裔和低收入人群产生不成比例的不利影响。此外,亨茨维尔地区 MPO 将为英语水平有限的人提供有意义的服务。

ADA 健康公平工具包和行动计划 2022-2025
作为普及和预防倡导委员会的成员,我们呼吁牙科界代表承认对所有人口腔健康的平等愿景。口腔健康是整体健康的重要组成部分。每个人都应享有基本的口腔健康,使每个人都能在健康状态下进食、工作、学习和生活,避免任何口腔疼痛或功能障碍。所有牙科安全网机构,无论是公共、私人还是两者的合作,都应提供全面的口腔保健服务,以满足个人和所在社区的需求。应适当考虑、设计和资助牙科保健服务,以满足社区的口腔健康需求,并充分扩大规模以发挥最大影响。口腔健康是任何医疗保健计划的重要组成部分,应考虑各级医疗保健专业人员所见的每位患者的持续健康和福祉。教育公众、立法者、媒体和教育界的努力应使人们认识到口腔健康的重要性,并感谢那些提供口腔健康预防护理和治疗服务的人。医疗保健计划应获得足够的资金,以提供与实践可行性相称的足够报销。慈善和志愿服务慷慨地满足了短期需求,因为它们减少了获得牙科护理的财务/交通障碍。我们可以通过开发更持久的解决方案来继续改善这些接入点,为所有人提供最优质的口腔保健。美国牙科协会呼吁口腔健康社区的所有方面,包括患者、倡导者、提供者和资助者,找到共同点,制定有效和可持续的口腔健康公平意识和实施战略。这种合作将促进疾病预防;减少医疗保健障碍,倡导对解决服务不足人群口腔健康需求的项目进行合理和充分的财政支持,并鼓励有效和高效的努力将口腔健康纳入整体健康。