XiaoMi-AI文件搜索系统
World File Search SystemAstrocytes
药房研究人员阐明了大脑如何处理触觉
Stobart补充说,触摸对象时人类在繁忙十字路口的高峰时段的流量就像高峰时段一样。“星形胶质细胞是将信息流向目的地的交通信号灯。当NMDA受体信号在星形胶质细胞中破坏时,就像左转的左转光一样。某些信息流可以通过交叉路口直接继续,但没有左转意味着某些信息无法达到目标。”

在小鼠中寻求
研究文章:新研究| Cognition and Behavior Astrocytes in the External Globus Pallidus Selectively Represent Routine Formation During Repeated Reward- Seeking in Mice https://doi.org/10.1523/ENEURO.0552-24.2025 Received: 3 December 2024 Revised: 3 February 2025 Accepted: 24 February 2025 Copyright © 2025 Yang et al.这是根据Creative Commons Attribution 4.0国际许可条款分发的开放访问文章,只要将原始工作正确归因于任何媒介,它允许在任何媒介中进行无限制的使用,分发和复制。

Göttingen会议德国神经科学学会2023Göttingen会议德国神经科学学会2023
1弗朗西大学,生物学跨学科研究中心,11个Place Marcelin Berthelot,75005,法国巴黎,我们的实验室研究了我们的实验室是否以及如何以及如何以及如何以及如何在信息处理中直接发挥了非常丰富的非神经元,但活性细胞的非常丰富的非神经元,但活跃的细胞。我们特别探讨了聚焦于离体或体内的生理和病理环境中星形胶质细胞 - 神经元相互作用的分子模态和功能结果。为此,我们使用一种多学科方法,结合了电生理学,成像,行为测试,数学建模和分子工具,将靶向选择性星形胶质细胞和小鼠和人体组织中的体内靶向。使用这种策略,我们在过去几年中进行了关于星形胶质细胞在正常和病理条件下突触传播,可塑性和网络活动中作用的基础研究。我们发现了调节生理和病理神经元活性的几种主要星形胶质细胞。,我们通过调节细胞外基质,离子稳态,胶质递质释放或星形胶质细胞突触覆盖范围来揭示连接素控制神经元接线和活性的多种方式。因此,我们的工作增添了神经网络的新兴概念,在该概念中,星形胶质细胞积极参与局部神经元网络的形成,活动和可塑性。

用...
摘要 - 传统的人工神经网络从生物网络中汲取灵感,使用神经元的节点层来传递信息进行处理。更现实的模型包括在神经网络中的尖峰,更贴近捕获电气特性。然而,很大一部分脑细胞是神经胶质细胞类型的,特别是星形胶质细胞被认为在执行计算中起作用。在这里,我们介绍了一个修改后的尖峰神经网络模型,并在神经网络中具有添加类似星形胶质细胞的单元,并评估它们对学习的影响。我们将网络作为液态机器实现,并任务网络执行混乱的时间序列预测任务。我们改变了网络中类似神经元和星形胶质细胞样单元的数量和比率,以检查后一种单元对学习的影响。我们表明,与神经和星形细胞网络相反,神经元和星形胶质细胞的结合对于推动学习至关重要。有趣的是,我们发现当类似星形胶质细胞样和神经元的单位之间的比率大约为2:1时,达到了最高的学习率,这反映了生物星形胶质细胞与神经元比率的一些估计值。我们的结果表明,在跨时间范围内代表信息的类似星形胶质细胞样单元可以改变神经网络的学习率,并且应将星形胶质细胞与神经元的比例适当地调整为给定的任务。

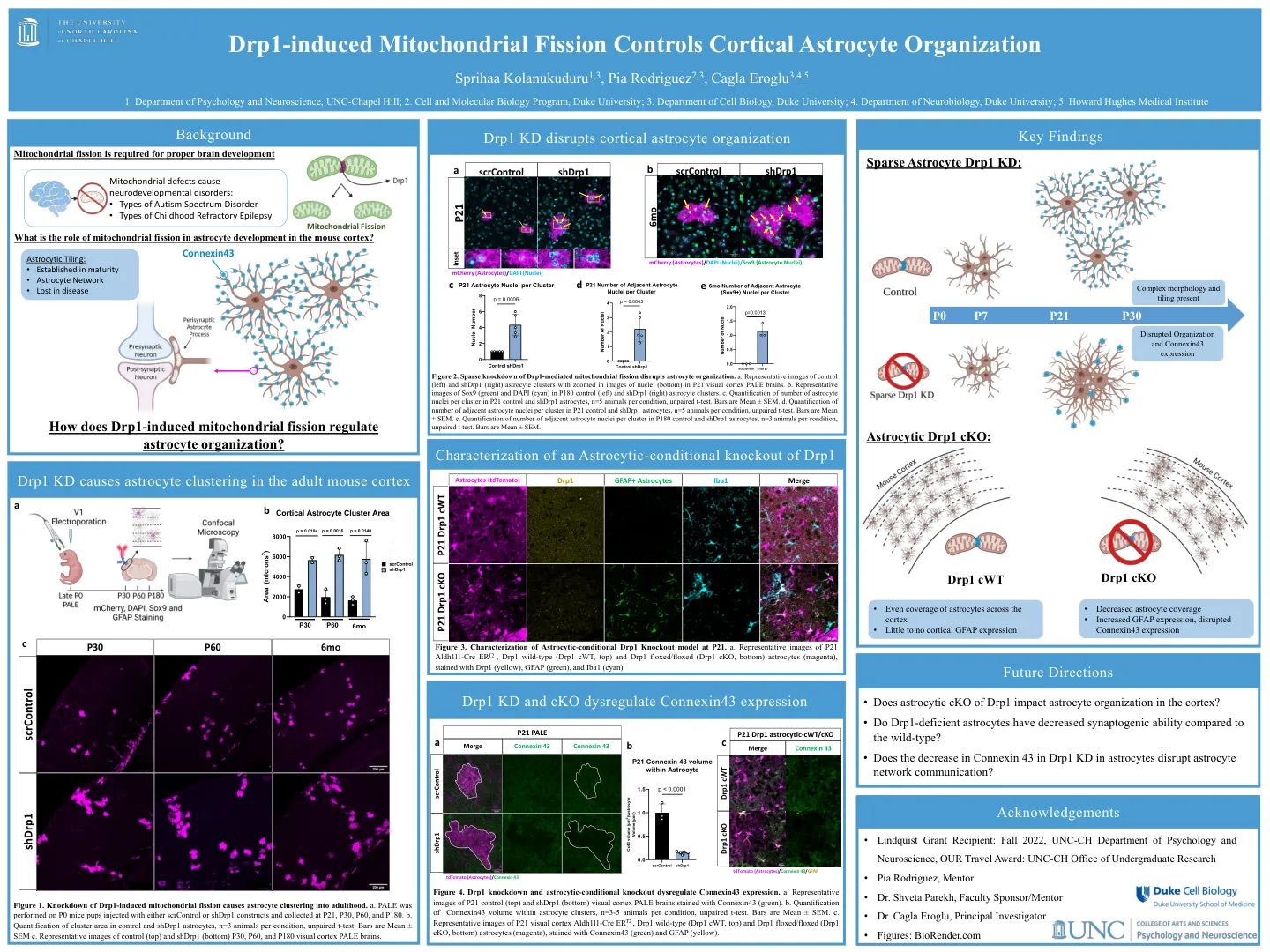
DRP1诱导的线粒体裂变控制皮质星形胶质细胞组织
图2。DRP1介导的线粒体裂变的稀疏敲低破坏了星形胶质细胞组织。a。对照(左)和SHDRP1(右)星形胶质细胞簇的代表性图像在p21视觉皮层中的核(底部)中放大。b。P180对照(左)和SHDRP1(右)星形胶质细胞簇中Sox9(绿色)和DAPI(青色)的代表性图像。c。 p21对照和SHDRP1星形胶质细胞中每个簇的星形胶质细胞核数量的量化,n = 5只动物,每个条件,未配对的t检验。条是平均值±SEM。d。 p21对照和SHDRP1星形胶质细胞中每个簇相邻星形胶质细胞核的数量,n = 5只动物,每个条件,未配对的t检验。条是平均值±SEM。e。 p180对照和SHDRP1星形胶质细胞中每个簇相邻星形胶质细胞核的数量定量,n = 3只动物,每个条件,未配对的t检验。条是平均值±SEM。

纹状体星形胶质细胞 A2A-D2 受体...
现在普遍认为星形胶质细胞是突触传递的活跃参与者,因此中枢神经系统中整合信号通讯的神经中心观点正在转向神经星形中心观点。星形胶质细胞对突触活动作出反应,释放化学信号(神经胶质递质)并表达神经递质受体(G 蛋白偶联受体和离子型受体),因此在中枢神经系统中充当神经元信号通讯的共同参与者。G 蛋白偶联受体通过异源化进行物理相互作用,形成具有新的独特信号识别和转导途径的异源体和受体嵌合体,这种能力在神经元质膜上得到了深入研究,并改变了中枢神经系统中整合信号通讯的观点。纹状体神经元质膜上的腺苷 A2A 和多巴胺 D2 受体是通过异源化进行受体间相互作用的最著名例子之一,对生理学和药理学观点都有相关影响。这里我们回顾了天然 A2A 和 D2 受体也可以通过星形胶质细胞质膜上的异源聚合相互作用的证据。发现星形胶质细胞 A2A-D2 异源聚合体能够控制纹状体星形胶质细胞突起释放谷氨酸。本文讨论了纹状体星形胶质细胞和星形胶质细胞突起上的 A2A-D2 异源聚合体在控制纹状体谷氨酸能传递方面的潜在相关性,包括在精神分裂症或帕金森病等病理条件下谷氨酸能传递失调的潜在作用。

基因
摘要:缺血性中风是全球严重发病率和死亡率的主要原因。中风后神经炎症最近受到了越来越多的关注,目的是为缺血性中风提供新的有效治疗策略。小胶质细胞和星形胶质细胞是中枢神经系统先天免疫系统的主要组成部分。他们可以参与IS-Chemic中风的所有阶段,从早期阶段,导致了第一波神经元细胞死亡,再到涉及吞噬作用和修复的晚期。在缺血性中风的早期阶段,小胶质细胞和星形胶质细胞的激活(通过星形胶质细胞连接素43半通道),触摸后造成神经炎性损伤之间存在恶性循环。然而,在缺血性中风的后期,反复活化的小胶质细胞可以通过触发侵入周围区域的反应性星形胶质细胞增多来诱导神经胶质疤痕的形成,这可能会限制反向激活的小胶质细胞运动,并限制炎症对健康组织的炎症扩散,使脑组织受伤,促进症状,从而受到刺激,并受到伤害。在这篇综述中,我们阐明了星形胶质细胞和小胶质细胞的各种作用,并总结了它们与神经炎症的关系。我们还研究了星形胶质细胞和小胶质细胞如何在缺血性中风的不同阶段相互影响。已经审查了靶向杂粒细胞和小胶质细胞的几种潜在的治疗方法。了解星形胶质细胞 - 小胶质细胞相互作用过程的细节将有助于更好地理解缺血性中风的机制,从而有助于鉴定新的治疗性干预措施。

审查对距离脊髓病变原发性遥控部位的细分市场的更多关注
多模式成像研究的最新发现表明,在脊髓和大脑中的脊髓损伤的震中,区域的宏观结构病理变化。正在进行研究以确定这些移位的细胞和分子机制,这些移位目前知之甚少。研究表明,重点区域中的病理过程是多方面的。此过程涉及星形胶质细胞和小胶质细胞,这有助于神经纤维从直接影响区域传播的神经纤维的变性,并参与相互激活。结果,距脊髓损伤位置的区域有突触损失。反应性星形胶质细胞产生硫酸软骨素蛋白聚糖,可抑制轴突生长和损伤细胞。但是,偏远地区的神经元死亡仍然有争议。原发性损伤面积是释放到脑脊液中的许多神经毒性分子的来源。假定这些分子(主要是基质金属蛋白酶)破坏了血脊髓屏障,从而导致偏远地区的巨噬细胞前体浸润。活化的巨噬细胞分泌促炎性细胞因子和基质金属蛋白酶,这反过来诱导了星形胶质细胞和小胶质细胞,一种促炎的表型。另外,反应性小胶质细胞与星形胶质细胞一起分泌了许多促炎和神经毒性分子,这些分子激活了炎症信号通路,从而加剧了突触耗竭和神经系统降解。似乎很可能是慢性炎症和神经退行性之间的相互作用是远离病变中心的脊髓区域中病理过程的关键特征。遥远地区的病理变化应成为潜在治疗靶标的研究对象。

神经元潮湿通过星形细胞RIPK3信号
引言最近的工作确定了神经炎症在神经系统疾病的发病机理中的核心作用,包括阿尔茨海默氏病和帕金森氏病等主要神经退行性疾病(1,2)。尽管神经胶质细胞是神经炎症的关键调节剂,但活化的神经胶质在疾病中起着复杂的作用,包括保护性和致病功能(3)。在神经胶质细胞中,星形胶质细胞是中枢神经系统(CNS)中最丰富的细胞类型,它们通过对神经传递,神经血管功能和代谢的广泛影响来支持稳态(4)。然而,在炎症性损伤后,星形胶质细胞可以进入与疾病发病机理相关的“反应性”状态(5)。尽管星形胶质细胞激活可能是高度塑性和背景依赖性的,但现在已广泛接受的是,星形胶质细胞可以在疾病期间采取炎症转录状态,这些疾病与神经毒性活性的赋予并抑制正常的稳态功能有关(6)。尽管有了这种理解,但在神经退行性疾病期间负责星形胶质细胞反应的分子机制,尤其是那些最直接加剧疾病进展的情况,仍然知之甚少(7)。