XiaoMi-AI文件搜索系统
World File Search SystemCCR5

检测异质性骨髓瘤患者中的 CCR5Δ32 突变等位基因...
CC 趋化因子受体 5 型 (CCR5 或 CD195) 是人类免疫缺陷病毒 (HIV) 的辅助受体结合位点之一。移植具有 CCR5 Δ 32 敲除突变的造血干细胞可以作为彻底治愈 HIV 的有效工具;这些方法已经通过了原理验证阶段。同时,使用现代 CRISPR/Cas9 基因组编辑方法,我们可以在任何野生型细胞中有效地复制 CCR5 Δ 32 突变。因此,在异质细胞混合物中寻找和准确量化突变 CCR5 Δ 32 等位基因的含量变得至关重要。在本研究中,我们描述了使用 CRISPR/Cas9 生成人工 CCR5 Δ 32 突变,然后进行多重液滴数字聚合酶链式反应 (ddPCR) 以量化其在细胞混合物中的含量。我们开发的系统可以让我们快速准确地测量 CCR5 Δ 32 突变细胞的含量,精确度可达 0.8%。

以 CCR5 为靶点,作为 HIV-1 治疗策略的一部分
在全球范围内,人类免疫缺陷病毒 1 型 (HIV-1) 感染是一种重大的健康负担,成功的治疗方案仍在研究中。目前已建立的终身抗逆转录病毒疗法 (ART) 的药物面临的挑战包括毒性、耐药 HIV-1 毒株的产生、治疗成本以及无法从受感染细胞中根除原病毒。出于这些原因,需要能够预防或消除疾病进展(包括获得性免疫缺陷综合征 (AIDS) 的发生)的新型抗 HIV-1 疗法。虽然 HIV-1 疫苗的开发也具有挑战性,但最近的进展表明,通过靶向 CC 趋化因子受体 5 型 (CCR5),可以预防 HIV-1 感染者感染 HIV-1 易感细胞。CCR5 在人类免疫反应中发挥多种功能,是 HIV-1 用来进入免疫细胞的辅助受体。针对 CCR5 的治疗通常涉及基因编辑技术,包括 CRISPR、使用抗体或拮抗剂阻断 CCR5,或两者结合。本文我们回顾了这些方法的有效性,并讨论了它们作为新型 ART 独立 HIV-1 感染疗法在临床中的应用潜力。

CCR5基因后进行后湿夫的阈值...
摘要自体,CCR5基因编辑的造血茎和祖细胞(HSPC)移植是实现HIV缓解的有前途的策略。但是,只能在体内编辑一小部分HSPC,以防止感染。要投影HIV缓解所需的CCR5版本的阈值,我们开发了一种数学模型,该模型概述了SHIV-1157IPD3N4感染的Pigailed Macaques的血液T细胞重建和血浆Simian-HIV(SHIV)动力学,该动力学与CCR5 Genee Editie ccr5 Gene Editied。该模型预测,当分析治疗中断(ATI)可以获得病毒控制时:(1)移植的HSPC至少比全身辐照后残留的内源性HSPC高至至少五倍,并且(2)受保护的HSPC的分数在移植中触发了阈值(76–94-94-94%),以使跨度的差异得出了跨度的差异,以胜过跨度的跨度。在这些条件下,如果扣留了ATI,直到移植基因改性细胞植入并重构为稳定状态,则预计会发生自发的病毒控制。

CRISPR-Cas9 诱导的 CCR5 Δ32 突变可作为抗 HIV-1 变体的有效基因治疗方法:综述
摘要。近五十年来,人类免疫缺陷病毒 (HIV) 一直是人类最大的流行病。尽管进行了广泛的研究,但迄今为止仍没有特定的治疗方法,CC 趋化因子受体 5,Delta 32 (CCR5 Δ32) 等位基因遗传点突变在预防获得性免疫缺陷综合征 (AIDS) 中起着至关重要的作用。这项综合研究旨在回顾在阳性患者群体中,在正选择压力下使用成簇的规律间隔短回文重复序列、Cas 9 酶 (CRISPR-Cas9) 和造血干细胞移植诱导纯合隐性缺失基因型作为表型。提出了一种方法,通过体细胞基因疗法,利用全能干细胞在受控的现代医疗设施内显著增加 Delta 32 有益突变等位基因的表达。它作用于两个功能失调的 CCR5 基因,翻译突变的 G 蛋白偶联共受体,其主要功能类似于 CXC 基序趋化因子受体 4 ( CXCR4 ),通过阻止病毒 RNA 进入 CD4 + T 辅助淋巴细胞,阻止感染并抑制病毒生命周期。这种修饰在北欧很常见,在那里它自然地以多态性 p (X=0.01) 的形式存在于高加索血统人群样本中,其中 X 是人群样本中对 HIV- 1 完全免疫的频率概率。分析了单核苷酸多态性 (SNP) 的表观遗传学,因为它们在免疫分布中起着重要作用。此外,在 CRIS-PR-Cas9 的伦理界限内进行了比较分析,以讨论所提出的方法和治疗替代方案的实际方面和挑战。此外,该研究汇集了所有可用数据并总结了先前的研究,同时为这一伦理困境提供了有希望的解决方案。最后,设计了一种方法来回答

靶向CCR5的新型TALEN的临床前评估证实了对HIV -1感染的抗性的功效和安全性
治疗感染人类免疫障碍病毒(HIV)的患者旨在防止病毒复制但无法消除病毒的疗法。尽管同种异体CCR5的移植32个纯合干细胞移植提供了治愈一些患者的治疗方法,但由于潜在的侧面影响,这种方法不被视为一般的治疗策略。相反,编码C-C趋化因子受体(CCR5)基因座的基因编辑(编码主要的HIV colecector)已显示出对CCR5-纤维化HIV菌株的抗性。在这里,提出了一种工程的转录激活剂样核酸酶(TALEN),可以在造血细胞中实现有效的CCR5编辑。将TALEN编码mRNA转移到原代CD4 + T细胞中后,高达89%的CCR5等位基因被破坏。基因分型证实了CCR5编辑的细胞的遗传稳定性,而全基因组的靶向分析确定了没有相关的诱变事件。在用CCR5-循环艾滋病毒挑战编辑的T细胞时,会观察到剂量依赖性的保护。功能评估表明,在增殖和它们在外源刺激时分泌细胞因子的能力方面,编辑和对照细胞之间没有显着差异。总而言之,一项高度活跃和特定的破坏CCR5已成功设计,为其在造血干细胞移植物中的临床应用铺平了道路。

引用本文为:sáez-cirión,A。等。野生型CCR5 Dono
这是接受出版的同行评审纸的PDF文件。尽管未经编辑,但内容已受到初步格式。自然医学正在为我们的作者和读者提供这本书的早期版本。文本和数字将在本文以最终形式发表之前进行复制和证明审查。请注意,在生产过程中可能会发现可能影响内容的错误,并且所有法律免责声明都适用。

通过调节NF-kB信号通路i,拮抗肽特异性结合CCR5,抑制炎症细胞浸润
引言:CC趋化因子受体5(CCR5)及NF-κB信号通路在炎症性肠病(IBD)的病理生理中起重要作用。前期我们合成了两条特异性与CCR5第一和第二个胞外环(分别为ECL1和ECL2)结合的多肽(GH肽和HY肽),并初步发现这两条肽对结肠炎有抑制作用。但这两条肽调控三硝基苯磺酸(TNBS)诱导的大鼠结肠炎的具体机制尚不清楚。本研究旨在进一步探讨CCR5结合肽在大鼠结肠炎中的作用及机制。材料与方法:用5%TNBS诱导实验性结肠炎。CCR5拮抗肽每天静脉注射一次,持续一周。通过组织学观察、实时定量PCR、Western印迹和相关性分析等方法观察CCR5结合肽对炎症细胞浸润和NF- κ B信号通路的影响。结果:给予GH和HY肽可减轻实验性结肠炎黏膜损伤,减少中性粒细胞、淋巴细胞和巨噬细胞的浸润(p < 0.05)。给予GH和HY肽后,NF- κ B相关基因p105、p100、IKK和TNF- α的表达降低(p < 0.01),TNF- α的蛋白水平以及IKK、I κ B α和p65的磷酸化也受到抑制。此外,CCR5拮抗肽可抑制p65的核转位。 Spear-man相关性分析显示炎症细胞的浸润与NF- κ B通路有显著相关性。结论:CCR5的ECL1和ECL2特异性结合拮抗肽通过调控NF- κ B信号通路抑制TNBS诱导的Sprague-Dawley大鼠结肠炎结肠黏膜中性粒细胞、淋巴细胞和巨噬细胞的浸润。

标题 经过基因编辑的 CD34+ 细胞源自人类 iPS 细胞(在体内但不是体外),可植入并分化为抗 HIV 细胞
iPS 细胞 | CCR5 | HIV 抗性 | 基因编辑 | 畸胎瘤 近期 HIV 研究的主要目标是开发一种“治愈”这种病毒感染的方法,避免终身接受抗逆转录病毒疗法 (ART)。实现这一目标的方法之一是删除或突变编码促进 HIV 感染和传播的蛋白质的基因。这一策略的一个有吸引力的候选基因是 Ccr5 基因,该基因突变导致 32 bp 缺失,已被证明与天然保护免受 HIV 感染和疾病有关 (1, 2)。Ccr5 基因编码 CCR5,这是一种人类细胞表面趋化因子受体,是 HIV 附着和感染细胞的辅助受体 (3, 4)。Ccr5 等位基因的 32 bp 缺失导致 CCR5 受体的截短异构体 CCR5 Δ 32,它不在细胞表面表达。因此,病毒进入细胞被阻止 (5)。诱导性多能干 (iPS) 细胞 (6) 能够分化为 CD34 + 造血干细胞 (HSC) (7),因此可以重建完整的免疫系统 (8, 9)。因此,这些 iPS 细胞是基因工程的首选目标。我们小组和其他小组已经证明,由健康个体 (10) 和接受 ART 治疗的 HIV 感染患者 (11) 的外周血单核细胞 (PBMC) 产生的 iPS 细胞可以经过基因编辑,使其 Ccr5 基因的野生型等位基因携带 Ccr5 Δ 32 突变 (12, 13)。值得注意的是,使用 CRISPR/Cas9 技术,可以修改 Ccr5 基因,使其具有与对 R5 嗜性病毒的抵抗力相关的天然 Δ 32 变体等位基因。此外,虽然截短的 CCR5 Δ 32 蛋白不存在于细胞表面,但它仍然表达,因此可能具有其他重要的生理作用(14-17)。我们已经证实,基因改造的 Ccr5 Δ 32 iPS 细胞可以在体外分化为 CD34 + HSC(10,18)。在适当的细胞培养条件下,它们可以产生各种
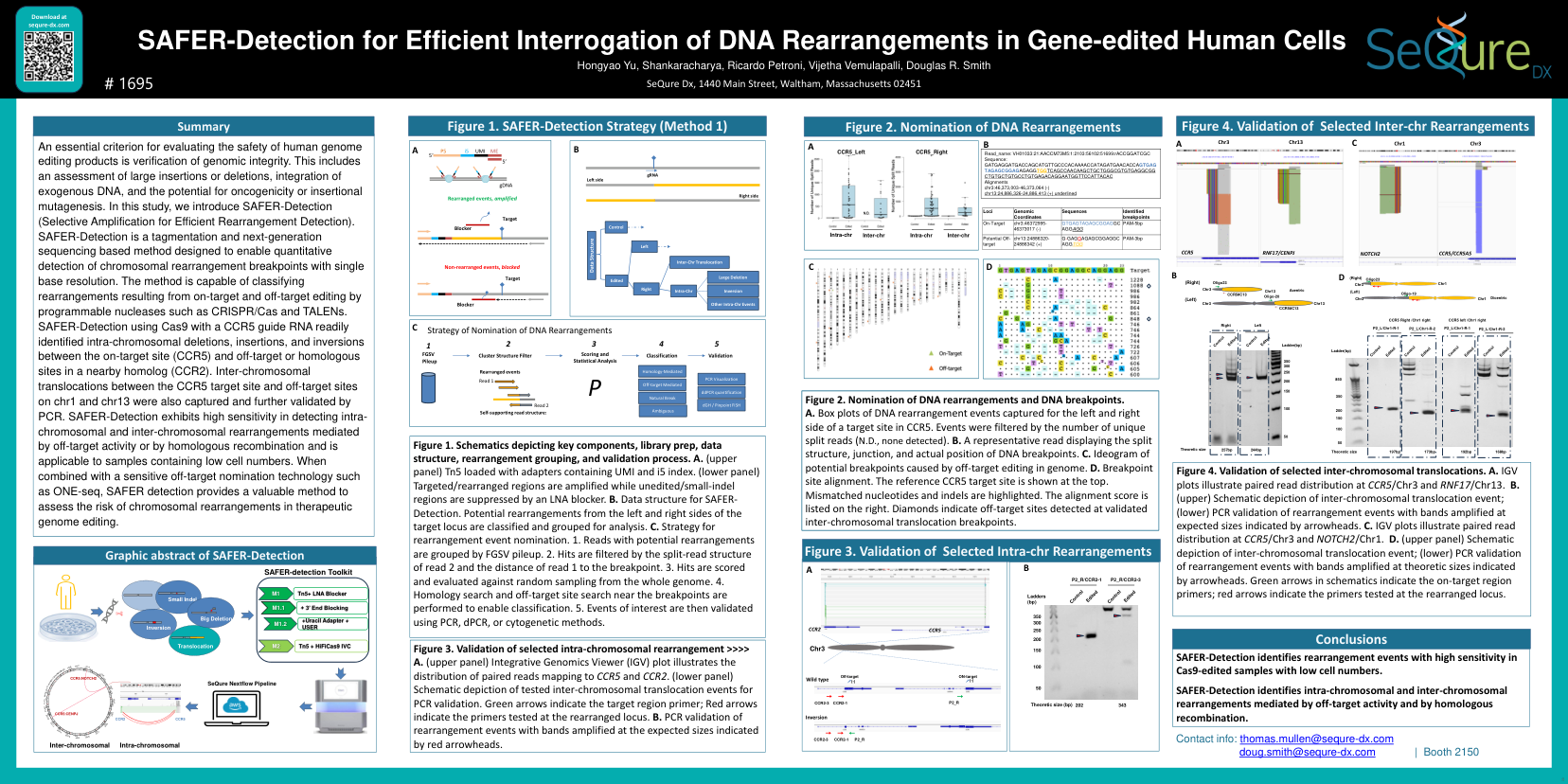
SAFER-Detection 用于有效检测基因编辑人类细胞中的 DNA 重排
评估人类基因组编辑产品安全性的一个重要标准是验证基因组完整性。这包括对大量插入或缺失、外源 DNA 整合以及致癌性或插入诱变可能性的评估。在本研究中,我们介绍了 SAFER-Detection(高效重排检测的选择性扩增)。SAFER-Detection 是一种基于标记和下一代测序的方法,旨在以单碱基分辨率定量检测染色体重排断点。该方法能够对由可编程核酸酶(如 CRISPR/Cas 和 TALEN)进行的靶向和脱靶编辑导致的重排进行分类。SAFER-Detection 使用 Cas9 和 CCR5 向导 RNA,可轻松识别靶位点 (CCR5) 与附近同源物 (CCR2) 中的脱靶或同源位点之间的染色体内缺失、插入和倒位。CCR5 靶位点与 chr1 和 chr13 上的脱靶位点之间的染色体间易位也被捕获并通过 PCR 进一步验证。SAFER-Detection 在检测由脱靶活动或同源重组介导的染色体内和染色体间重排方面表现出高灵敏度,适用于含有低细胞数的样本。当与灵敏的脱靶提名技术(如 ONE-seq)结合使用时,SAFER 检测提供了一种评估治疗性基因组编辑中染色体重排风险的宝贵方法。

穿梭肽在体内将碱基编辑器 RNP 递送至恒河猴气道上皮细胞
我们在此报告了首次证明穿梭肽在恒河猴模型中将蛋白质和 ABE8e-Cas9 RNP 递送至呼吸道上皮的转化潜力。在单次气雾剂给药后,我们成功地将荧光标记的蛋白质货物递送至大气道和小气道的上皮细胞以及一些肺泡上皮。使用 S315 穿梭肽进行 ABE8e-Cas9 RNP 递送,我们在使用支气管刷回收的细胞中实现了 CCR5 基因座的显著 A 到 G 编辑。从气管和近端气道收获的上皮中 CCR5 位点的编辑效率达到 5.3%。在具有 R553X 突变的人类 CF 气道上皮中应用这种递送方法实现了类似的编辑水平并赋予 CFTR 功能的部分恢复。