XiaoMi-AI文件搜索系统
World File Search SystemCCR5
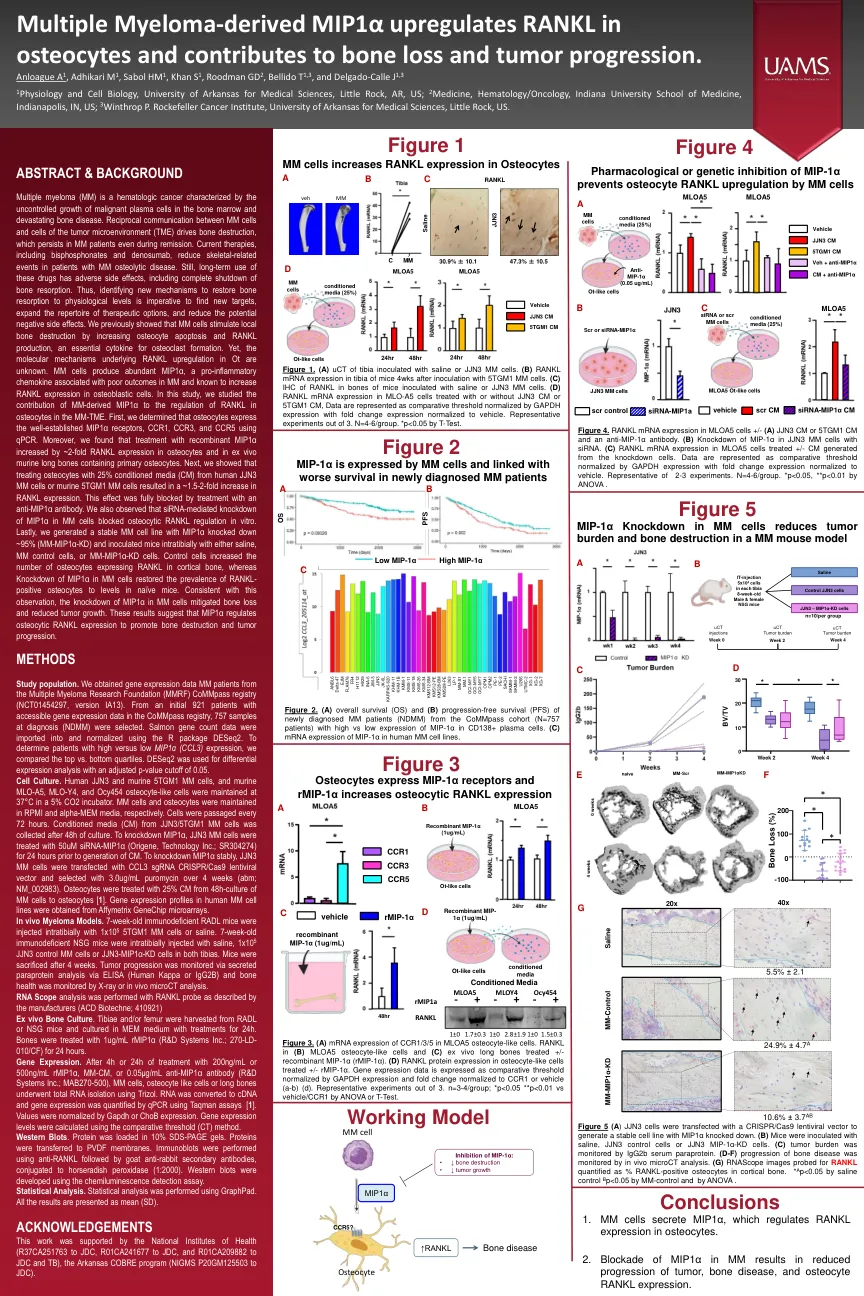
多发性骨髓瘤的MIP1α上调RANKL ...
多发性骨髓瘤(MM)是一种血液学癌,其特征是骨髓和毁灭性骨病中恶性血浆细胞的生长不受控制。MM细胞与肿瘤微环境(TME)细胞之间的相互通信驱动骨骼破坏,即使在缓解过程中,MM患者也持续存在。当前的疗法,包括双膦酸盐和denosumab,减少了MM骨化疾病患者的骨骼相关事件。仍然,这些药物的长期使用具有不利的副作用,包括骨吸收的完全关闭。因此,必须确定新的机制以将骨吸收恢复到生理水平,这对于寻找新靶标,扩展治疗选择的曲目并减少潜在的负面副作用至关重要。我们先前表明,MM细胞通过增加骨细胞凋亡和RANKL产生来刺激局部骨破坏,这是破骨细胞形成的必不可少的细胞因子。然而,OT中RANKL上调的分子机制尚不清楚。mM细胞产生丰富的MIP1α,这是一种与MM结局不佳相关的促炎性趋化因子,并且已知会增加成骨细胞中RANKL表达。在这项研究中,我们研究了MM衍生的MIP1α对MM-TME中骨细胞中RANKL调节的贡献。首先,我们确定骨细胞使用qPCR表达了良好的MIP1α受体CCR1,CCR3和CCR5。此外,我们发现重组MIP1α的治疗在骨细胞和含有原发性骨细胞的离体鼠长骨中增加了约2倍的RANKL表达。接下来,我们表明,从人JJN3 MM细胞或鼠5TGM1 MM细胞中使用25%条件培养基(CM)处理骨细胞,RANKL表达增加了约1.5-2倍。用抗MIP1α抗体处理完全阻断了这种作用。我们还观察到siRNA介导的MM细胞中MIP1α的敲低阻断了体外骨细胞RANKL调节。最后,我们生成了一个稳定的MM细胞系,MIP1α击倒了〜95%(MM-MIP1α-KD),并用盐水,MM对照细胞或MM-MIP1α-KD细胞在室内接种小鼠。对照细胞增加了在皮质骨中表达RANKL的骨细胞的数量,而MM细胞中MIP1α的敲低使RANKL-阳性骨细胞的患病率恢复为幼稚小鼠的水平。与该观察结果一致,MM细胞中MIP1α的敲低减轻骨质流失和肿瘤生长减少。这些结果表明MIP1α调节骨细胞RANKL表达以促进骨骼破坏和肿瘤进展。
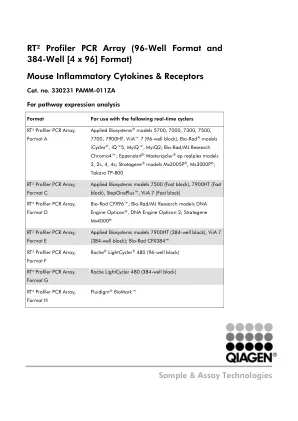
RT² Profiler PCR 阵列(96 孔格式和 384 ... - GeneGlobe
A01 Mm.235137 NM_007926 Aimp1 氨酰 tRNA 合成酶复合物相互作用多功能蛋白 1 A02 Mm.103205 NM_007553 Bmp2 骨形态发生蛋白 2 A03 Mm.1283 NM_011329 Ccl1 趋化因子(CC 基序)配体 1 A04 Mm.4686 NM_011330 Ccl11 趋化因子(CC 基序)配体 11 A05 Mm.867 NM_011331 Ccl12 趋化因子(CC 基序)配体 12 A06 Mm.41988 NM_011332 Ccl17 趋化因子(CC 基序)配体 17 A07 Mm.424740 NM_011888 Ccl19 趋化因子(CC 基序)配体 19 A08 Mm.290320 NM_011333 Ccl2 趋化因子(CC 基序)配体 2 A09 Mm.116739 NM_016960 Ccl20 趋化因子(CC 基序)配体 20 A10 Mm.12895 NM_009137 Ccl22 趋化因子(CC 基序)配体 22 A11 Mm.31505 NM_019577 Ccl24 趋化因子(CC 基序)配体 24 A12 Mm.1282 NM_011337 Ccl3 趋化因子(CC 基序)配体 3 B01 Mm.244263 NM_013652 Ccl4 趋化因子(CC 基序)配体 4 B02 Mm.284248 NM_013653 Ccl5 趋化因子(CC 基序)配体 5 B03 Mm.137 NM_009139 Ccl6 趋化因子(CC 基序)配体 6 B04 Mm.341574 NM_013654 Ccl7 趋化因子(CC 基序)配体 7 B05 Mm.42029 NM_021443 Ccl8 趋化因子(CC 基序)配体 8 B06 Mm.416125 NM_011338 Ccl9 趋化因子(CC 基序)配体 9 B07 Mm.274927 NM_009912 Ccr1 趋化因子(CC 基序) 受体 1 B08 Mm.8021 NM_007721 Ccr10 趋化因子 (CC 基序) 受体 10 B09 Mm.6272 NM_009915 Ccr2 趋化因子 (CC 基序) 受体 2 B10 Mm.57050 NM_009914 Ccr3 趋化因子 (CC 基序) 受体 3 B11 Mm.1337 NM_009916 Ccr4 趋化因子 (CC 基序) 受体 4 B12 Mm.14302 NM_009917 Ccr5 趋化因子 (CC 基序) 受体 5 C01 Mm.8007 NM_009835 Ccr6 趋化因子 (CC 基序) 受体 6 C02 Mm.442098 NM_007720 Ccr8 趋化因子(CC 基序)受体 8 C03 Mm.4861 NM_011616 Cd40lg CD40 配体 C04 Mm.795 NM_007778 Csf1 集落刺激因子 1(巨噬细胞) C05 Mm.4922 NM_009969 Csf2 集落刺激因子 2(粒细胞-巨噬细胞) C06 Mm.1238 NM_009971 Csf3 集落刺激因子 3(粒细胞) C07 Mm.103711 NM_009142 Cx3cl1 趋化因子(C-X3-C 基序)配体 1 C08 Mm.21013 NM_008176 Cxcl1 趋化因子(CXC 基序)配体 1 C09 Mm.877 NM_021274 Cxcl10 趋化因子(CXC 基序)配体 10 C10 Mm.131723 NM_019494 Cxcl11 趋化因子(CXC 基序)配体 11 C11 Mm.303231 NM_021704 Cxcl12 趋化因子(CXC 基序)配体 12 C12 Mm.10116 NM_018866 Cxcl13 趋化因子(CXC 基序)配体 13 D01 Mm.64326 NM_011339 Cxcl15 趋化因子(CXC 基序)配体 15 D02 Mm.4660 NM_009141 Cxcl5 趋化因子(CXC 基序)配体 5 D03 Mm.766 NM_008599 Cxcl9 趋化因子(CXC 基序)配体 9 D04 Mm.234466 NM_009909 Cxcr2 趋化因子(CXC 基序)受体 2 D05 Mm.12876 NM_009910 Cxcr3 趋化因子(CXC 基序)受体 3 D06 Mm.6246 NM_007551 Cxcr5 趋化因子(CXC 基序)受体 5 D07 Mm.3355 NM_010177 Fasl Fas 配体(TNF 超家族,成员 6) D08 Mm.240327 NM_008337 Ifng 干扰素伽马 D09 Mm.379327 NM_008348 Il10ra 白细胞介素10 受体,α

UCSF
越来越多的证据表明,产生白介素17(IL-17)的CD4 T细胞广泛称为T辅助17(Th17)细胞,有助于控制结核病。在MTB感染的青少年中,IL-17的转录特征降低了与非培训者相比,那些发展为活动性结核病的人的血液降低(1);另一项研究发现,对MTB抗原产生IL-17的CD4 T细胞的子集比非进展者中的MTB抗原不那么丰富(2)。MTB响应性CD4 T细胞在人肺中富集,并且与血浆IL-1 B成反比,这表明在MTB的控制中起作用(3)。il-17 - 产生的CD4 T细胞在人免疫去毒(HIV)感染中被耗尽,其耗竭有助于通过(4,5)中的肠粘膜屏障功能分解,在(4、5)中逐渐取决于CCR5的表达,从而有助于辅助症状。使用几种方法来识别T辅助17个细胞(本文表示为“ Th17”细胞,包括所有子集,而不论所使用的标准如何)。刺激时产生的 IL-17的产生是Th17细胞的典型特征(6,7)。 尽管该鉴定了Th17细胞,但并未对Th17谱系中的所有细胞进行分类,而其他细胞可以产生IL-17(8-11)。 在健康供体的血液和类风湿关节炎患者的滑膜流体中,CCR6 +子集包含所有IL-17 - 产生表达RORC mRNA的T细胞(12),这是支持人类Th17分化的转录因子。 宿主代谢会影响免疫反应(24),包括通过驱动细胞分化。IL-17的产生是Th17细胞的典型特征(6,7)。尽管该鉴定了Th17细胞,但并未对Th17谱系中的所有细胞进行分类,而其他细胞可以产生IL-17(8-11)。在健康供体的血液和类风湿关节炎患者的滑膜流体中,CCR6 +子集包含所有IL-17 - 产生表达RORC mRNA的T细胞(12),这是支持人类Th17分化的转录因子。宿主代谢会影响免疫反应(24),包括通过驱动细胞分化。使用CCR6作为Th17细胞的标志物已在幼稚的脐带血(13),炎症性疾病(14)中复制(14),以及TB等感染(15)。细胞表面标记CD26(16-19)和CD161(20-23)也已用于鉴定Th17细胞。CD26 HI CD4 + T细胞被证明是共表达CD161和CCR6,并富含生产IL-17(16)。在转录组研究中,通过RORC的表达鉴定Th17细胞IL23R(支持IL-17 - 产生细胞的扩张)和IL-17 mRNA。一个例子是色氨酸分解代谢的Kynurenine途径,其产品(共同称为Kynurenines)在促进Treg发育时抑制了Th17(25 - 28)。在结核病中,潜在结核感染(LTBI)的人循环色氨酸(TRP)浓度下降,随着TB治疗而发展为主动TB(29)和TRP浓度增加(25)。同样,感染HIV的人具有低循环的TRP浓度,并与发病机理联系起来(30)。血浆kynurenine/色氨酸比率(K/T)可以将活性结核病的人与没有HIV共感染的多药耐药(MDR-TB)患者中的人体区分开,并且与对照组相比,多药抗药性(MDR-TB)患者的人数明显更高(25)。血浆K/T在艾滋病毒(PWH)的患者中也升高,尤其是在那些发展为艾滋病的人中(27,31)。尽管在HIV中建立了K/T与Treg/Th17比率之间的链接,但在TB中缺乏类似的数据。在这里,我们表征了由ART抑制的HIV(HIV-ART)在参与者中定义的Th17细胞子集,以确定IL-17 - 产生对MTB抗原反应的CD4 T细胞是否在HIV-ART中改变并测试