XiaoMi-AI文件搜索系统
World File Search SystemCCR8

教练-CCR8、TEAD1 和 HER2
2023 年 12 月,COACH 召开了第七次会议,与美国国家癌症研究所 (NCI)、美国食品药品管理局 (FDA)、欧洲药品管理局 (EMA)、倡导团体、制药行业、儿科临床前概念验证平台 (ITCC-P4) 和儿科临床前体内测试 (PIVOT) 联盟的代表会面,以评估三个癌症相关目标,这些目标可能是开发用于儿科适应症的救命药物的潜在实体。

趋化因子受体靶向治疗:CCR8 的特殊情况
摘要:针对细胞毒性 T 淋巴细胞相关蛋白 4 (CTLA-4) 和程序性死亡受体 1 (PD-1) 或其配体 1 (PD-L1) 的免疫检查点阻断抑制剂 (CBI) 改变了许多癌症患者的前景。从转化的角度来看,这一显著进展凸显了免疫细胞在控制肿瘤进展中的重要性。仍有改进空间,因为目前的 CBI 疗法只使少数患者受益。此外,干扰免疫检查点受体经常会导致免疫相关不良事件 (irAE),并在某些患者中产生危及生命的后果。肿瘤微环境 (TME) 中的免疫抑制细胞,包括肿瘤内调节性 T (Treg) 细胞、肿瘤相关巨噬细胞 (TAM) 和髓源性抑制细胞 (MDSC),会促进肿瘤进展并与不良疾病前景相关。最近的报告显示趋化因子受体 CCR8 在肿瘤 Treg 细胞上有选择性表达,这使得 CCR8 成为转化研究中有希望的靶点。在这篇综述中,我总结了我们目前对 CCR8 在生理和病理生理过程中的细胞分布和功能的了解。讨论包括评估去除表达 CCR8 的细胞可能如何影响抗肿瘤免疫以及远程位点的免疫稳态。基于这些考虑,CCR8 似乎是未来转化研究中值得考虑的一个有希望的新靶点。

通过常规T细胞获得抑制功能,限制了Treg Depettion
调节性T(T Reg)细胞有助于免疫稳态,但抑制了对癌症的免疫反应。破坏T Reg细胞介导的癌症免疫抑制的策略已达到有限的临床成功,但是对治疗衰竭的非衍生机制知之甚少。通过对小鼠的T reg细胞靶向免疫疗法进行建模,我们发现CD4 + FOXP3-常规T(T Conv)细胞在耗尽Foxp3 + T Reg细胞时获得抑制功能,从而限制了治疗功效。foxp3 -t conver细胞在消融T reg细胞时采用T型细胞样转录曲线,并获得抑制T细胞激活和增殖的能力。sup-压力活性富含。在T型细胞耗竭后,CCR8 + T CORS细胞会经历全身性和肿瘤内激活和扩张,并介导IL-10-依赖性抑制抗肿瘤免疫。因此,T细胞内IL10的有条件缺失增强了小鼠T型细胞耗竭后的抗肿瘤免疫力,而IL-10信号传导的抗体阻断随着T reg细胞的耗竭协同以克服治疗耐药性。这些发现揭示了T Conv细胞在治疗性T型细胞耗竭后释放的T Cons细胞的二级免疫抑制层,并表明在T细胞谱系中更广泛考虑抑制功能以开发有效的T reg toR taR剂量剂量的治疗疗法。
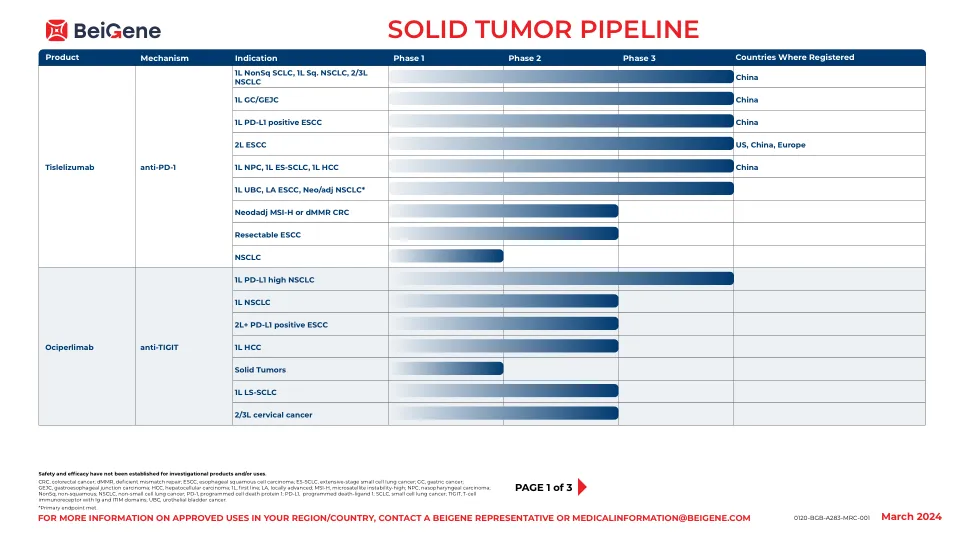
实体瘤管道
TNBC,三阴性乳腺癌; CCR8,CC趋化因子受体8; CD3,分化3群; CRC,结直肠癌; CSF-1R,刺激性因子-1受体; DGKζ,二酰基甘油激酶ζ; DKK1,Dickkopf-1; DLL3,类似三角洲的配体3; ESCC,食管鳞状细胞癌; FGFR,成纤维细胞生长因子受体; GBRCAM,种系BRCA突变; GC-GEJC,胃癌 - 糖食管癌; HCC,肝细胞癌; 1L,第一行; MCRPC,转移性cast割前列腺癌; MTX,维护处理; NSCLC,非小细胞肺癌; PARP,聚(ADP-核糖)聚合酶; PI3KΔ,磷酸肌醇3-激酶三角洲; PSOC,铂敏感的卵巢癌; RAF,快速加速的纤维肉瘤; STEAP1,前列腺1的六跨膜上皮抗原; VEGFR,血管内皮生长因子受体。
简介方法结果
缩写:1L,第一行; 2L,第二行; BC,乳腺癌; BSAB,双特异性抗体; BTC,胆道癌; CCR8,C-C基序趋化因子受体8; CRC,结直肠癌; DGKζ,二酰基甘油激酶ζ; GC,胃癌; GEA,胃食管腺癌; HER2,人表皮生长因子受体2; HNSCC,头和颈部鳞状细胞癌HPK1,造血祖细胞激酶1; lag3,淋巴细胞激活基因3; LS-SCLC,有限阶段的小细胞肺癌; MBC,转移性乳腺癌; MSS,微卫星稳定性; OX40,肿瘤坏死因子(TNF)受体家族的成员,也称为CD134。 RCC,肾细胞癌; r/r,复发/耐火; SCLC,小细胞肺癌; SMAC,第二个线粒体衍生的caspase激活剂; TIM3,T细胞免疫球蛋白结构域和粘蛋白结构域3; UC,尿路上皮癌。

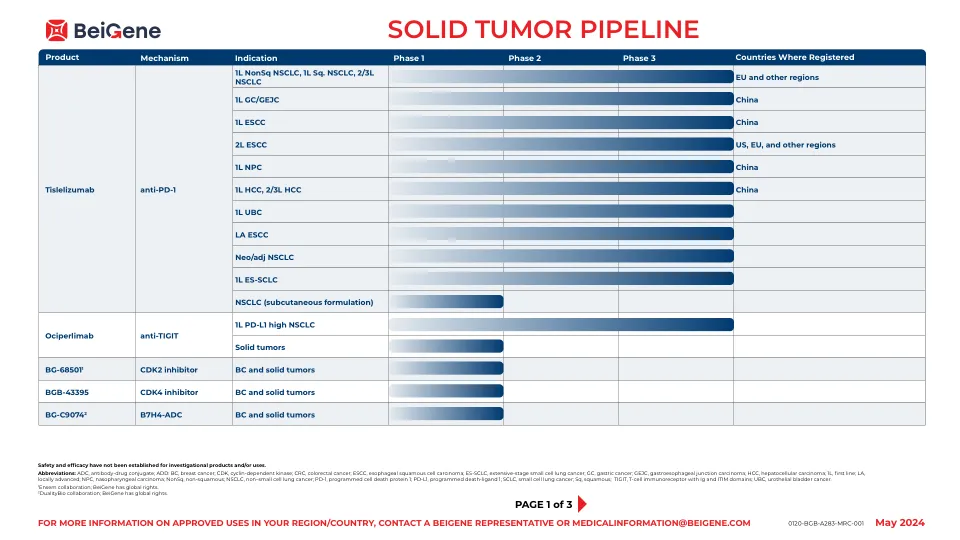
实体瘤管道
缩写:1L,第一行; 2L,第二行; 3L,第三行; 4-1BB,肿瘤坏死因子受体超家族成员9; ADC,抗体 - 药物结合; BSAB,双特异性抗体; CCR8,C-C基序趋化因子受体8; CD16A,FC受体FCγRIIIA; CD3,分化3群; CDAC,嵌合降解激活化合物; CEA,癌脑抗原; DGKζ,二酰基甘油激酶ζ; DLL3,类似三角洲的配体3; EGFR,表皮生长因子受体; ES-SCLC,广泛的小细胞肺癌; FGFR2B,成纤维细胞生长因子受体2,同工型IIIB; GBRCAM,种系乳腺癌基因(BRCA)突变;胃肠道,胃肠道; GPC3,Glypican-3; IL-15,白介素15; Kras,Kirsten大鼠肉瘤病毒; LS-SCLC,有限阶段的小细胞肺癌; MCRPC,转移性cast割前列腺癌; MTX,维护处理; MUC1,粘蛋白1; PARP 1/2,聚(ADP-核糖)聚合酶1和2; PRMT5,蛋白精氨酸甲基转移酶5; PSOC,铂敏感的卵巢癌; SCLC,小细胞肺癌; STEAP1,前列腺1的六跨膜上皮抗原。

文章表达重组的改性疫苗病毒Ankara通过重编程Tregs
在肿瘤微环境中,免疫抑制调节细胞(TREG)的有效耗竭而不触发全身自身免疫性是癌症免疫疗法的重要策略。改性疫苗Ankara(MVA)是一种高度减弱的非复制疫苗病毒,具有悠久的人类使用史。在这里,我们报告了免疫激活重组MVA(RMVA,MVAδE5R-FLT3L-OX40L),其vacinia e5r基因的缺失(编码DNA传感器cyclic cyclice cgas,cgas,cgas的抑制剂),cgas和cgas的抑制剂,cgas和表达3个抑制剂) OX40L。肿瘤内(IT)RMVA(MVAδE5R-FLT3L-OX40L)产生有效的抗肿瘤免疫力,取决于CD8 + T细胞,CGAS/STING介导的介导的细胞溶质性DNA传感途径和I型I IFN信号。值得注意的是,它通过OX40L/OX40的相互作用和IFNAR信号传导来耗尽OX40 HI调节T细胞OX40 HI调节T细胞。用RMVA处理的肿瘤的单细胞RNA-SEQ分析显示OX40 HI CCR8 HI tregs的耗竭以及IFN反应性Tregs的膨胀。综上所述,我们的研究提供了通过免疫激活RMVA耗尽和重编程的肿瘤内Treg的概念证明。

2024; 15(3): 632-644. doi: 10.7150/jca.89581 研究论文探索头颈部鳞状细胞癌的预后免疫微环境相关基因
目的:头颈部鳞状细胞癌 (HNSCC) 具有较高的局部和远处转移率。在肿瘤组织中,肿瘤细胞与肿瘤微环境 (TME) 之间的相互作用与癌症的发展和预后密切相关。因此,筛选 HNSCC 中的 TME 相关基因对于了解转移模式至关重要。方法:我们的研究主要依赖于一种名为使用表达数据估计恶性肿瘤中的基质和免疫细胞 (ESTIMATE) 的新算法。从 TCGA 数据库中获取外显子模型每百万映射片段的每千碱基片段 (FPKM) 数据和 HNSCC 临床数据,并确定 HNSCC 组织的纯度以及基质和免疫细胞浸润的特征。此外,根据免疫、基质和 ESTIMATE 评分筛选差异表达基因 (DEG),并评估它们的蛋白质-蛋白质相互作用 (PPI) 网络和 ClueGO 功能。最后,确定了 HNSCC 中与免疫相关的 DEG 的表达谱。在高侵袭性口腔癌细胞系 (SCC-25、CAL-27 和 FaDu) 和口腔癌组织中验证了差异基因表达。结果:我们的分析发现免疫和 ESTIMATE 评分均与 HNSCC 的预后显着相关。此外,使用 Venn 算法进行交叉验证显示 433 个基因显着上调,394 个基因显着下调。所有 DEG 都与 ESTIMATE 和免疫评分相关。使用通路富集分析观察到细胞因子-细胞因子受体相互作用和趋化因子信号通路的富集。在分析了 PPI 网络的关键子网络后,我们初步筛选了 25 个基因。生存分析揭示了 CCR4、CXCR3、P2RY14、CCR2、CCR8 和 CCL19 与生存的关系以及它们与 HNSCC 中免疫浸润相关转移的关系。结论:使用 ESTIMATE 对基质和免疫细胞进行评分后,筛选了相关 TME 相关基因的表达谱,并确定了与生存相关的 DEG。这些 TME 相关基因标记物可作为 HNSCC 中的预后指标和指示转移性状的标记物,具有宝贵的实用性。
RT² Profiler PCR 阵列(96 孔格式和 384 ... - GeneGlobe
A01 Mm.235137 NM_007926 Aimp1 氨酰 tRNA 合成酶复合物相互作用多功能蛋白 1 A02 Mm.103205 NM_007553 Bmp2 骨形态发生蛋白 2 A03 Mm.1283 NM_011329 Ccl1 趋化因子(CC 基序)配体 1 A04 Mm.4686 NM_011330 Ccl11 趋化因子(CC 基序)配体 11 A05 Mm.867 NM_011331 Ccl12 趋化因子(CC 基序)配体 12 A06 Mm.41988 NM_011332 Ccl17 趋化因子(CC 基序)配体 17 A07 Mm.424740 NM_011888 Ccl19 趋化因子(CC 基序)配体 19 A08 Mm.290320 NM_011333 Ccl2 趋化因子(CC 基序)配体 2 A09 Mm.116739 NM_016960 Ccl20 趋化因子(CC 基序)配体 20 A10 Mm.12895 NM_009137 Ccl22 趋化因子(CC 基序)配体 22 A11 Mm.31505 NM_019577 Ccl24 趋化因子(CC 基序)配体 24 A12 Mm.1282 NM_011337 Ccl3 趋化因子(CC 基序)配体 3 B01 Mm.244263 NM_013652 Ccl4 趋化因子(CC 基序)配体 4 B02 Mm.284248 NM_013653 Ccl5 趋化因子(CC 基序)配体 5 B03 Mm.137 NM_009139 Ccl6 趋化因子(CC 基序)配体 6 B04 Mm.341574 NM_013654 Ccl7 趋化因子(CC 基序)配体 7 B05 Mm.42029 NM_021443 Ccl8 趋化因子(CC 基序)配体 8 B06 Mm.416125 NM_011338 Ccl9 趋化因子(CC 基序)配体 9 B07 Mm.274927 NM_009912 Ccr1 趋化因子(CC 基序) 受体 1 B08 Mm.8021 NM_007721 Ccr10 趋化因子 (CC 基序) 受体 10 B09 Mm.6272 NM_009915 Ccr2 趋化因子 (CC 基序) 受体 2 B10 Mm.57050 NM_009914 Ccr3 趋化因子 (CC 基序) 受体 3 B11 Mm.1337 NM_009916 Ccr4 趋化因子 (CC 基序) 受体 4 B12 Mm.14302 NM_009917 Ccr5 趋化因子 (CC 基序) 受体 5 C01 Mm.8007 NM_009835 Ccr6 趋化因子 (CC 基序) 受体 6 C02 Mm.442098 NM_007720 Ccr8 趋化因子(CC 基序)受体 8 C03 Mm.4861 NM_011616 Cd40lg CD40 配体 C04 Mm.795 NM_007778 Csf1 集落刺激因子 1(巨噬细胞) C05 Mm.4922 NM_009969 Csf2 集落刺激因子 2(粒细胞-巨噬细胞) C06 Mm.1238 NM_009971 Csf3 集落刺激因子 3(粒细胞) C07 Mm.103711 NM_009142 Cx3cl1 趋化因子(C-X3-C 基序)配体 1 C08 Mm.21013 NM_008176 Cxcl1 趋化因子(CXC 基序)配体 1 C09 Mm.877 NM_021274 Cxcl10 趋化因子(CXC 基序)配体 10 C10 Mm.131723 NM_019494 Cxcl11 趋化因子(CXC 基序)配体 11 C11 Mm.303231 NM_021704 Cxcl12 趋化因子(CXC 基序)配体 12 C12 Mm.10116 NM_018866 Cxcl13 趋化因子(CXC 基序)配体 13 D01 Mm.64326 NM_011339 Cxcl15 趋化因子(CXC 基序)配体 15 D02 Mm.4660 NM_009141 Cxcl5 趋化因子(CXC 基序)配体 5 D03 Mm.766 NM_008599 Cxcl9 趋化因子(CXC 基序)配体 9 D04 Mm.234466 NM_009909 Cxcr2 趋化因子(CXC 基序)受体 2 D05 Mm.12876 NM_009910 Cxcr3 趋化因子(CXC 基序)受体 3 D06 Mm.6246 NM_007551 Cxcr5 趋化因子(CXC 基序)受体 5 D07 Mm.3355 NM_010177 Fasl Fas 配体(TNF 超家族,成员 6) D08 Mm.240327 NM_008337 Ifng 干扰素伽马 D09 Mm.379327 NM_008348 Il10ra 白细胞介素10 受体,α