XiaoMi-AI文件搜索系统
World File Search SystemCRBN

自旋 - 纬度放松的化学控制,以发现室温分子值
方式。然后将活化的泛素部分通过转化反应转移到泛素结合酶(E2)中。接下来,E2酶与多种泛素连接酶(E3)合作,将泛素转移至底物蛋白上的赖氨酸残基。多泛素化的底物蛋白被某些蛋白质识别,展开和降解。蛋白质降解器是小分子化合物,通过劫持细胞内UPS诱导靶蛋白的选择性降解。这种靶向蛋白质降解策略极大地扩展了可药物靶蛋白的光谱,并被认为是药物开发的一种新的有希望的方式,因为它不需要在靶蛋白上明确的活性位点。沙利度胺衍生物,例如列纳奈度胺和pomali-Di-Dimide,统称为免疫调节性伊酰亚胺药物(IMIDS),主要用于治疗血液性恶性肿瘤,例如多发性骨髓瘤(图2)。多发性骨髓瘤是一种疾病,其中骨髓中的浆细胞变为癌。 IMID和其他药物的开发


第56届中西部学生生物医学研究论坛
鉴定沙利度胺,多纳拉替胺和莫利度胺作为脑(CRBN)粘合剂刺激了促蛋白水解靶向嵌合体(Protacs)的发展。带有CRBN结合部分的PROTAC是与CRBN形成三元复合物的异功能分子,它募集了CUL4-DDB1泛素连接酶蛋白蛋白复合物和感兴趣的蛋白质(POI),并靶向其降解。尽管进行了许多持续的努力,但基于结构的Protac设计仍然难以捉摸。当前的方法依赖于Protac库的合成并以所选测定格式筛选库来识别可行的命中,这可能是耗时的和资源密集的,强调了对更有效的筛选方法的需求。为了应对这一挑战,我们假设使用HCT116野生型(WT)POI敲除(POI - / - )和CRBN敲除(CRBN - / - )细胞的基于生存力的表型筛选系统将能够快速识别POI和CRBN依赖性方式以POI和CRBN依赖性方式显示活性。在这项研究中,我们选择了MAP3K1(一种致癌信号激活剂),作为我们的POI并使用各种接头将ATP竞争力的MAP3K1抑制剂与Thalidomide共轭,以生成库。使用上述WT,MAP3K1 - / - 和CRBN - / - 单元线对库进行3天的生存能力屏幕。屏幕将50-008识别为有选择性抑制WT细胞的生存能力而不是MAP3K1 - / - 或CRBN - / - 细胞的命中。50-008在结直肠癌细胞系和类器官模型中进行了化学验证,从而产生了影响CRBN结合,MAP3K1结合或接头的单个变化。随访研究表明,50-008抑制了癌细胞的增殖,诱导凋亡,并以CRBN和MAP3K1依赖性方式抑制了S期细胞周期停滞。此外,在约900个癌细胞系中,在50-008的混合物(PRISM)筛选中同时进行了相对抑制,鉴定出267个敏感的癌细胞系(IC 50 <0.1 µM),包括腔内B乳腺癌细胞系T47D和131个抗性癌细胞系(IC 50> 10 µM)。在T47D细胞中的机理研究表明,50-008降解MAP3K1并调节关键下游效应子(例如ERK和IKKβ)的激活。总而言之,我们的表型筛选系统,我们确定并验证了50-008是一种有效的MAP3K1靶向Protac,具有多种癌症模型的选择性活性。这种方法建立了一种快速可靠的管道,用于识别功能性protac,同时验证其细胞特异性和作用机理。我们的发现不仅为未来的Protac开发提供了验证的筛选平台,而且还为靶向MAP3K1依赖性癌症提供了治疗潜力。

药理学和毒理学年度评论从沙利度胺到用于靶向蛋白质降解的合理分子胶设计
沙利度胺及其衍生物是强效的癌症治疗药物,也是最容易理解的分子胶降解剂 (MGD) 之一。这些药物选择性地重新编程 E3 泛素连接酶 cereblon (CRBN),使靶蛋白被泛素-蛋白酶体系统降解。MGD 在 E3 连接酶表面产生新的识别界面,参与诱导的蛋白质-蛋白质与新底物的相互作用。对其作用机制的分子洞察为通过特定的识别基序 G 环与大量靶标进行接触提供了令人兴奋的机会。我们的分析表明,目前基于 CRBN 的 MGD 原则上可以识别人类蛋白质组中超过 2,500 种包含 G 环的蛋白质。我们回顾了在调整 CRBN 与其 MGD 诱导的新底物之间的特异性方面的最新进展,并推断出一组控制这些相互作用的简单规则。我们得出结论,合理的 MGD 设计工作将能够选择性降解更多的蛋白质,从而将这种治疗方式扩展到更多的疾病领域。

一种评估E3连接酶用于靶向蛋白质降解
抽象评估靶蛋白降解(TPD)的潜在〜700 E3连接酶的适用性的主要挑战之一是缺乏针对每个E3连接酶的粘合剂。在这里,我们将遗传密码扩展(GCE)用于编码含四嗪的非典型氨基酸(TET-NCAA)位点特定于E3连接酶,可以通过在活着的细胞中与新的蛋白质蛋白质培养细胞一起将其连接到新的植物蛋白质蛋白培养细胞中。可以用Neo-Substrate的TPD评估所得的E3连接酶最小化和功能化的最小化和功能化。我们证明,用可单击的TET-NCAA编码的CRBN可以在已知的免疫调节药物(IMID)中或跨表面编码,可以共价连接到STCO-LINKER-JQ1和招募BRD2/4的crbn介导的降解,以表明CRBN的高塑料tpd。降解效率取决于在CRBN上编码的TET-NCAA的位置以及接头的长度,显示了这种方法在绘制E3连接酶表面识别最佳TPD口袋的能力。这种Elef-脱脂剂的方法不仅具有维持E3连接酶的天然状态,而且还允许在细胞内条件下对E3连接酶和靶蛋白伴侣进行询问,并且可以应用于任何已知的E3连接酶。关键字:泛素 - 蛋白酶体系统,靶向蛋白质降解,E3连接酶,Cereblon,遗传代码扩展,四嗪单击化学

作者索引
LBA002 Targeting GSPT1 by a novel cereblon E3 ligase modulator for the treatment of Acute Lymphoblastic Leukemia Fatemeh Keramatnia 1 , Yunchao Chang 1 , Gisele Nishiguchi 1 , Jaeki Min 1 , Charles Mullighan 1 , Marcus Fischer 1 , Zoran Rankovic 1 , Fatemeh Keramatnia 1 .1田纳西州孟菲斯的圣裘德儿童研究医院。急性淋巴细胞白血病(所有),最常见的儿童癌症和成人第二常见的急性白血病,是由骨髓中未分化的淋巴前体细胞的克隆扩张引起的。尽管大多数儿童期所有病例都具有转录因子(TF)基因突变或重排的克隆遗传改变,但TF改变仍然是困难的治疗靶标。小分子诱导的蛋白质降解是一种新型策略,可以应用于当前不受限制的靶标,例如TF和融合癌蛋白。在此范式中,小分子降解器(Protac或Mocular Glue(MG))重定向细胞的内源性泛素蛋白酶体系统,并诱导靶蛋白或非本地蛋白质的泛素化或非本地底物E3依基酶(Neosubstrate)(Neosubstrate)(Neosubstrate)及其下层蛋白酶质量下生成。最近,据报道,CRBN E3连接酶调节剂CC90009在急性髓细胞性白血病中表现出有效的抗肿瘤活性,从而导致GSPT1(G1至S相变因子)为CRBN Neosubstrate。这些发现表明,MGS针对不同恶性肿瘤中意外漏洞的潜力。我们在一组代表性急性白血病细胞系中对MGL进行筛选,包括CRLF2-重新排列的所有细胞系MHH-CALL-4鉴定了几个活性MG,具有EC50 <5μm。在这里,使用确认的CRBN结合亲和力,使用结构上多样化和独特的MGS(Molecular Glue库(MGL)),我们试图通过表型和蛋白质组学方法鉴定新的CRBN调节剂。Lenalidomide竞争分析和MHH-CALL-4 CRBN击倒细胞证实了这些MGS的CRBN依赖性。在这些化合物中,SJ6986,沙利度胺驱动的磺酰胺在10多个在体外测试的所有细胞系中显示出有效的细胞毒性。TMT-MS蛋白质组学分析将GSPT1/2鉴定为具有高选择性的该化合物的主要靶标。我们接下来在一组衍生的异种移植物(PDX)中测试了SJ6986的活性,该患者具有IGH-CRLF2,EPOR,ATF7IP-JAK2 EX VIVO的重排。所有测试的肿瘤对IC50在纳摩尔范围内的SJ6986高度敏感。NSG小鼠中的PK分析表明SJ6986的迅速吸收和超过80%的口服生物利用度。PD研究在IGH-CRLF2 PDX中显示出治疗后48小时内GSPT1的剂量依赖性降解。 最后,我们检查了6种不同的PDX中SJ6986的抗肿瘤活性,代表所有人的高风险亚型,包括近单倍体,低h-高dip虫,CRLF2重键和重排,在体内28天。 在大多数肿瘤模型中, SJ6986能够以1 mg/kg剂量大大减轻肿瘤负担。 共同确认,SJ6986是一种新型的CRBN调节剂和潜在的治疗剂,它通过靶向具有高选择性和效力的GSPT1蛋白来治疗所有人。PD研究在IGH-CRLF2 PDX中显示出治疗后48小时内GSPT1的剂量依赖性降解。最后,我们检查了6种不同的PDX中SJ6986的抗肿瘤活性,代表所有人的高风险亚型,包括近单倍体,低h-高dip虫,CRLF2重键和重排,在体内28天。SJ6986能够以1 mg/kg剂量大大减轻肿瘤负担。共同确认,SJ6986是一种新型的CRBN调节剂和潜在的治疗剂,它通过靶向具有高选择性和效力的GSPT1蛋白来治疗所有人。

PROTAC 降解剂临床试验总结
2010年,Itoh等人利用甲基乌苯美司合成了另一种PROTAC分子,以募集E3连接酶(凋亡蛋白抑制剂(IAP))来降解POI。为了提高效力和靶标选择性,具有高亲和力和特异性的小分子(例如,募集E3连接酶cereblon(CRBN)的邻苯二甲酰亚胺或识别E3连接酶Von Hippel-Lindau(VHL)的VHL-1)进入PROTAC分子,进而下调多种癌症靶标,例如Ikaros家族锌指蛋白1/3(IKZF1/3)和雌激素相关受体α(ERRα)。基于小分子的PROTAC的突破为PROTAC作为癌症治疗策略开辟了一条新道路。

分子胶的进展:探索靶向蛋白质降解的化学空间和设计原理
E3 连接酶 cereblon (CRBN) 被发现是沙利度胺及其类似物的靶标,这彻底改变了靶向蛋白质降解 (TPD) 领域。这种泛素介导的降解途径首先由二价降解剂利用。最近,低分子量分子胶降解剂 (MGD) 的出现扩大了 TPD 领域,因为 MGD 通过相同的机制运作,同时提供与小分子疗法一致的有吸引力的物理化学特性。本综述深入探讨了 MGD 的发现和发展,并以细胞周期蛋白 K 和锌指蛋白 IKZF2 为例进行了研究,重点介绍了设计原理、生物测定和治疗应用。此外,它还研究了分子胶的化学空间,并概述了推动该领域创新的合作努力。
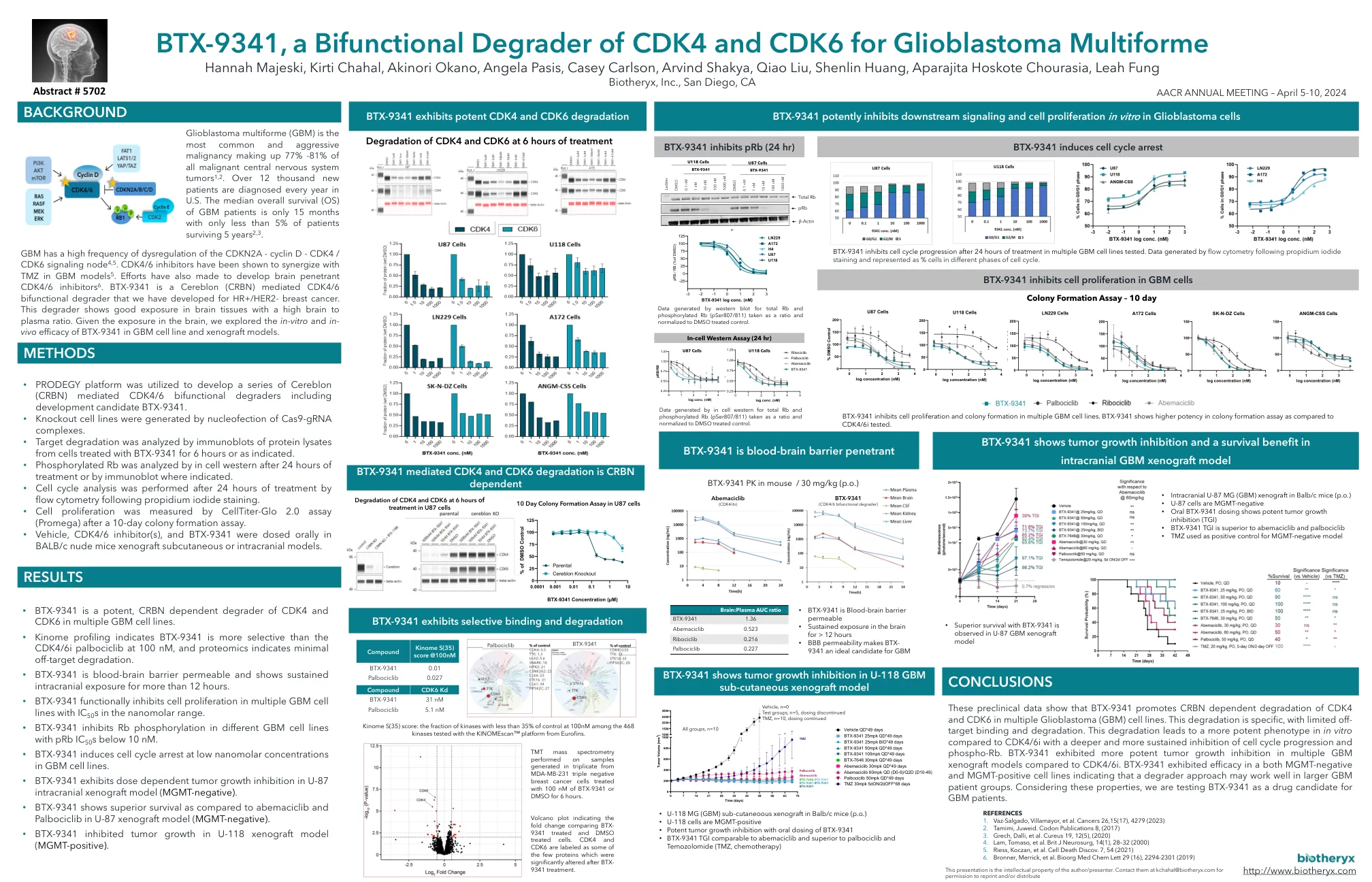
BTX-9341,CDK4和CDK6的双功能降解器,用于多形胶质母细胞瘤
•Prodegy平台用于开发一系列少数(CRBN)介导的CDK4/6双功能降解器,包括开发候选BTX-9341。•通过cas9-grna复合物的核反射产生基因敲除细胞系。•通过用BTX-9341处理的细胞的蛋白质裂解物的免疫印迹分析了靶降解6小时或所示。•经过24小时的治疗后或通过指示的免疫印迹,通过细胞西部分析了磷酸化的RB。•在碘化丙啶染色后通过流式细胞仪治疗24小时后,进行了细胞周期分析。•通过10天菌落形成测定后,通过CellTiter-Glo 2.0分析(Promega)测量细胞增殖。•媒介物,CDK4/6抑制剂和BTX-9341在BALB/C裸鼠异种皮下或颅内模型中口服。

mllt1/3 的首创靶向蛋白质降解剂,用于
通过 HTRF 测定法测量 MLLT1/3 YEATS 域抑制。除非另有说明,实验均在 MV4:11 细胞中进行。在 4 小时时确定人类 MLLT1 的降解。在 NIH-3T3 细胞中,在 5 小时时确定小鼠 MLLT1 和 3 的降解。使用 100nM MLLT-TPD 进行降解动力学分析。使用 JESS Protein Simple 确定 DC 50 和动力学。DIA 质谱全局蛋白质组学用于评估 10nM (4 小时) MLLT-TPD 的选择性。硼替佐米用作蛋白酶体抑制剂,来那度胺用作 CRBN 粘合剂。MLLT-I 是一种内部专有的 MLLT1/3 抑制剂,与 MLLT-TPD 密切相关。通过 Cell-TiterGlo 读数 (5d) 在 Elplasia 板中测量 AML/ALL 细胞活力。 MLLT-TPD 用于除染色质 MLLT1 降解(接近 MLLT-TPD 类似物,DC 50 1.4nM)和 AML/ALL 细胞活力(第二个接近 MLLT-TPD 类似物,DC 50 10nM)之外的所有实验。