XiaoMi-AI文件搜索系统
World File Search SystemCREB

失活肺癌对异常 CRTC- CREB 激活的影响
1 美国佛罗里达大学医学院分子遗传学与微生物学系;2 美国佛罗里达大学健康癌症中心;3 美国佛罗里达大学医学院生物化学与分子生物学系;4 美国佛罗里达大学遗传学研究所;5 中国广州中山大学中山眼科中心国家眼科学重点实验室;6 中国海宁浙江大学医学院国际校区浙江大学-爱丁堡大学研究所(ZJU-UoE Institute);7 美国三角研究园国家环境健康科学研究所(NIEHS)生殖与发育生物学实验室;8 美国佛罗里达大学医学院医学系

cAMP–PKA–CREB信号在癌症中的复杂作用
摘要 环磷酸腺苷 (cAMP) 是第一个发现的第二信使,在细胞信号转导中起着关键作用,调节许多生理和病理过程。cAMP 可以调节各种靶基因的转录,主要通过蛋白激酶 A (PKA) 及其下游效应物如 cAMP 反应元件结合蛋白 (CREB)。此外,PKA 可以磷酸化许多激酶,如 Raf、GSK3 和 FAK。异常的 cAMP-PKA 信号转导与各种类型的人类肿瘤有关。特别是,cAMP 信号转导可能具有肿瘤抑制和肿瘤促进作用,具体取决于肿瘤类型和环境。cAMP-PKA 信号转导可以调节癌细胞的生长、迁移、侵袭和代谢。本综述重点介绍了 cAMP-PKA-CREB 信号在肿瘤发生中的重要作用。还讨论了针对该途径进行癌症治疗的潜在策略。关键词:cAMP、PKA、CREB、癌症

ZDHHC16 通过抑制脑卒中模型中的 CREB 促进神经细胞铁死亡
引言:本研究探讨了ZDHHC16在脑卒中(CA)模型中的作用及其可能的机制。材料和方法:从我院收集CA患者。使用小鼠建立大脑中动脉闭塞(MCAO)模型。结果:CA患者的ZDHHC16水平上调。ZDHHC16上调在体外模型中促进炎症并加速线粒体损伤。ZDHHC16基因上调促进神经细胞铁死亡。抑制ZDHHC16可预防小鼠脑卒中。ZDHHC16上调通过促进CREB泛素化与CREB相互连接来抑制CREB。CREB激动剂抑制了体外模型中ZDHHC16上调的影响。 CREB 抑制剂在体外模型中抑制了 ZDHHC16 下调的影响。结论:我们得出结论,ZDHHC16 通过抑制 CREB 促进 CA 模型中的铁死亡和炎症。该发现可能对 CA 或其他神经系统疾病的治疗有益。

阿尔茨海默氏病的氧化应激
摘要阿尔茨海默氏病的病理生理学仍然是一个难题。越来越多的证据阐明了氧化应激参与AD的病理学,使其成为治疗性发育的主要靶标。由线粒体功能改变,电子传输链失调和其他来源产生的活性氧(ROS)提升了凝集的Aβ和神经原纤维缠结,从而进一步刺激了ROS的产生。氧化应激引起对脂质,蛋白质和DNA的损伤导致神经元死亡,从而导致AD。此外,氧化应激会诱导凋亡,这是由ERK1/2和NRF2途径的调节触发的,随后GSK-3β表达增加并降低了PP2A活性。氧化应激通过干扰RCAN1,CREB/ ERK,NRF2,PP2A,NFκB和PI3K/ AKT等各种信号通路来夸大疾病状况。研究报道了TNF-α在氧化应激刺激中的作用,该抗氧化剂刺激的作用增强了抗氧化剂水平。据报道,其他药物如普拉己烯,美金刚,卡维丝醇和褪黑激素可以激活CREB/RCAN1和NRF2途径。与此相一致,epigallocatechin Gallate和Genastein还靶向NRF2和CREB途径,从而导致下游途径(如AS和KEAP1)的激活,这些途径可以改善氧化应激条件。多奈酮和白藜芦醇减少氧化应激,并激活AMPK途径以及PP2A激活,从而促进tau去磷酸化和神经元存活。本研究详细描述了氧化应激在AD中的作用,涉及氧化应激诱导的AD的主要信号通路和正在针对这些途径的开发中的药物,这些途径可能有助于AD的治疗进展。

INCB160058的临床前评估
AKT,蛋白激酶B; CREB,环状腺苷单磷酸反应元件结合蛋白;细胞仪,飞行时间的细胞仪; DMSO,二甲基磺氧化物; ERK,细胞外信号调节激酶; IRF,干扰素调节因素; Jak,Janus激酶; MAPKAPK,有丝分裂原激活的蛋白激酶激活的蛋白激酶; MEK,有丝分裂原激活的蛋白激酶激酶; MTOR,雷帕霉素的哺乳动物靶标; PI3K,磷酸肌醇-3激酶; STAT,信号换能器和转录激活因子; TPO,血小子蛋白; wt,野生型。AKT,蛋白激酶B; CREB,环状腺苷单磷酸反应元件结合蛋白;细胞仪,飞行时间的细胞仪; DMSO,二甲基磺氧化物; ERK,细胞外信号调节激酶; IRF,干扰素调节因素; Jak,Janus激酶; MAPKAPK,有丝分裂原激活的蛋白激酶激活的蛋白激酶; MEK,有丝分裂原激活的蛋白激酶激酶; MTOR,雷帕霉素的哺乳动物靶标; PI3K,磷酸肌醇-3激酶; STAT,信号换能器和转录激活因子; TPO,血小子蛋白; wt,野生型。

爱泼斯坦 - 巴尔病毒,维生素D和免疫反应
2型糖尿病(T2D)和肥胖症是影响全球数百万个人的普遍代谢性疾病。一种称为Tirzepatide治疗肥胖和T2D的新有效的治疗药物是GIP受体和GLP-1受体的双重激动剂。tirzepatide在临床上比GLP-1受体激动剂更有效,但原因尚未得到充分理解。tirzepatide比GLP-1受体更有效地刺激GIP受体。然而,尚未在E354(Wildtype)或Q354(E354Q)GIP受体变体上进行彻底研究Tirzepatide信号传导。E354Q变体与T2D和低体重指数的风险相关。为了更好地了解GIP受体信号传导,我们表征了两个GIP受体变体中内源性激动剂和Tirzepatide的活性。使用COS7细胞,我们检查了WildType和E354Q GIP受体信号传导,分析CAMP和IP 1的积累以及AKT,ERK1/2和CREB磷酸化。GIP(1-42)和GIP(1-30)NH 2在不包括CREB磷酸化的这些途径上显示出等值效应,其中GIP(1-30)NH 2在E354Q GIP受体上比GIP(1-42)更有效。tirzepatide在两个变体中都偏爱cAMP信号。这些发现表明Tirzepatide是一种偏向GαS信号的激动剂,并表明它同样激活了WildType和E354Q GIP受体变体。我们还观察到具有内源性肽的GIP受体变体的药理学之间的差异,这可能有助于解释表型的差异。这些发现有助于对GIP受体信号的全面理解,并将有助于开发打击T2D和肥胖症的疗法。

2021; 12(13): 4099-4108. doi: 10.7150/jca.58097 研究论文 长链非编码RNA FIRRE促进肝细胞增殖和糖酵解
最近的研究表明,长链非编码RNA (lncRNA) FIRRE 参与结直肠癌和弥漫大B细胞淋巴瘤的增殖、抗凋亡和侵袭。然而,FIRRE 在肝细胞癌 (HCC) 中的生物学功能仍然未知。在本文中,我们发现与非肿瘤组织相比,HCC 中的 FIRRE 水平经常升高。与正常肝细胞相比,我们还证实了 HCC 细胞中 FIRRE 水平上调。值得注意的是,FIRRE 高表达与恶性临床特征有关,包括晚期 TNM 分期和肿瘤大小≥5 cm,并导致 HCC 生存率较低。功能上,FIRRE 敲低抑制了 HCCLM3 细胞的增殖和糖酵解。FIRRE 过表达增强了 Huh7 细胞增殖和糖酵解。值得注意的是,FIRRE 正向调节肝癌细胞中的乙醇酸酶 6-磷酸果糖-2-激酶/果糖-2,6-双磷酸酶 4 (PFKFB4) 表达。PFKFB4 在肝癌组织中高表达且与 FIRRE 水平呈正相关。PFKFB4 的上调表达与高肿瘤分级和晚期 TNM 分期相关。TCGA 数据显示,PFKFB4 高表达提示肝癌预后不良。从机制上讲,调节 FIRRE 水平不会影响 PFKFB4 mRNA 的稳定性。FIRRE 主要分布在肝癌细胞的细胞核中,并通过 cAMP 反应元件结合蛋白 (CREB) 促进 PFKFB4 的转录和表达。PFKFB4 可以消除 FIRRE 敲低对肝癌细胞增殖和糖酵解的影响。总之,高表达的 FIRRE 通过增强 CREB 介导的 PFKFB4 转录和表达促进 HCC 细胞增殖和糖酵解。

癌细胞国际诱导基因表达
图1创建合成cAMP响应元件结合蛋白(CREB)响应启动子。(a)腺苷信号传导的描述。腺苷(红色球)结合腺苷受体A2AR/A2BR,该腺苷受体动员相关的G蛋白(绿色)激活腺苷酸环化酶(橙色受体),并将ATP转化为3'5'- 5'-循环腺苷单磷酸腺苷(Camp)。另外,福斯科蛋白(橙色球)可以直接激活腺苷循环酶。CAMP结合蛋白激酶A(PKA)与磷酸化的CREB,该CREB结合了Plindromic DNA基序“ TGACGTCA”,激活了基因表达。(b)启动子设计和筛选示意图。cAMP响应元件基序(CRE,突出显示的黄色)被克隆在3倍重复中,两侧是鸟嘌呤“ G”(带下划线),六个散布的填充核苷酸(N)。3x Cres(灰色正方形)放在核心启动子(蓝色箭头)上游的1-6个重复中。用高斯荧光素酶(GLUC)或绿色荧光蛋白(EGFP)定量启动子活性。(c,d)HEK293T细胞在96个井板中用指示的构建体(x轴)反向转染。转染后48小时,用车辆(DMSO,浅蓝色条)或20μm福斯科林(FSK,深蓝色条)将细胞介质更改为培养基。八个小时后,对培养基进行了采样并测试了GLUC活性(RLU)。条表示n = 3实验重复的平均值,误差线代表标准误差(SEM)。**通过方差分析(ANOVA)Tukey检验,与所有其他样本相比,表示P <0.01。(E,F)流式细胞仪启动子诱导。HEK293T细胞用96个井板中的指定构建体(x轴)反向转染。转染后48小时,细胞培养基被更改为未处理的培养基(浅蓝色条),或补充了0.750 m m m腺苷(ADO,深蓝色条)的培养基。八个小时后,将细胞胰蛋白酶胰蛋白酶进行胰蛋白酶,并将其重悬于FACS缓冲液中以进行流式细胞仪。y轴表示正向散射(FSC)单元的EGFP中位荧光强度。条代表n = 3实验重复的平均值,误差线代表SEM。(g)启动子对腺苷的剂量反应性。HEK293T细胞在96个井板上反向转染,并在传说中指示的构造,然后培养48小时。然后更改培养基以添加不同的腺苷浓度,在8小时后进行采样,并测试了GLUC活性(RLU)。**通过12倍-CRE_YB的ANOVA TUKEY测试代表P <0.01,与1 m m的所有其他样品相比。每个点表示n = 3实验重复的平均值,误差线为SEM。
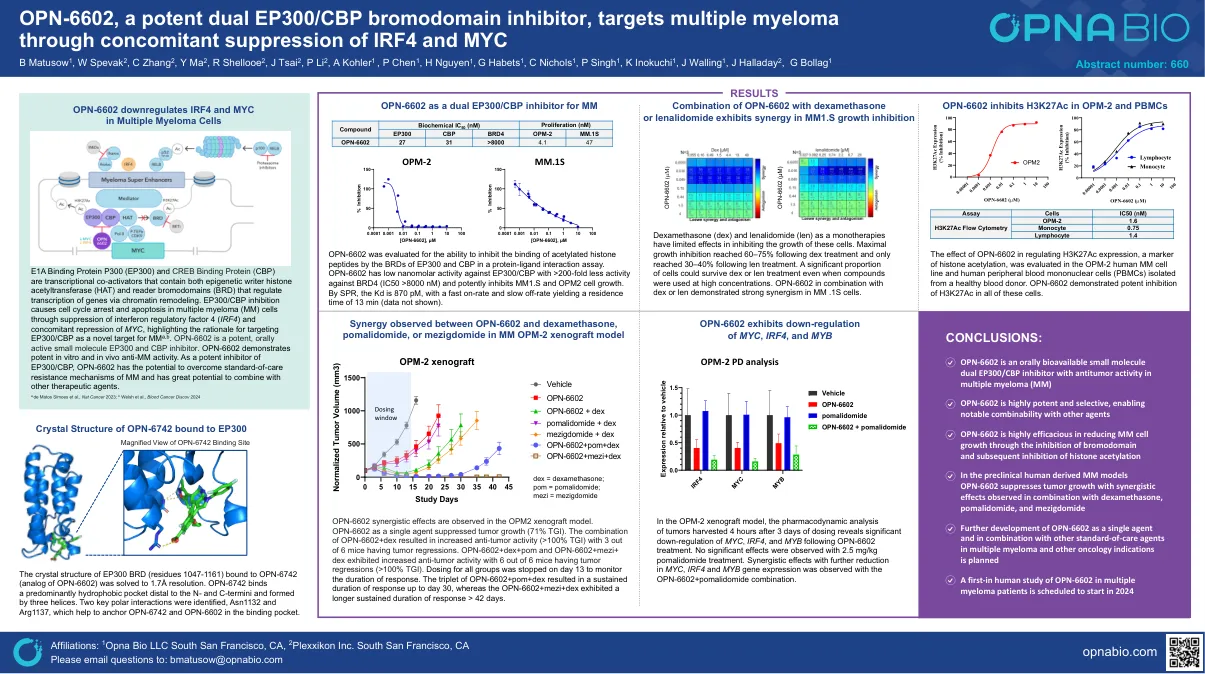
OPN-6602,一种有效的双EP300/CBP溴化域抑制剂,通过同时抑制IRF4和MYC
e1a结合蛋白p300(EP300)和CREB结合蛋白(CBP)是转录共激活因子,既包含表观遗传作者组蛋白乙酰基转移酶(HAT)和读取子溴化酶(BRD),并且通过染色质重塑调节基因的转录。EP300/CBP抑制作用通过抑制干扰素调节因子4(IRF4)和伴随抑制MYC导致多发性骨髓瘤(MM)细胞的细胞周期停滞和凋亡,并强调将EP300/CBP作为MM A,b,b,b。OPN-6602是一种有效的,有效的小分子EP300和CBP抑制剂。OPN-6602表现出体外和体内抗MM活性的有效性。 作为EP300/CBP的有效抑制剂,OPN-6602具有克服MM的护理标准抗性机制的潜力,并且具有与其他治疗剂相结合的巨大潜力。OPN-6602表现出体外和体内抗MM活性的有效性。作为EP300/CBP的有效抑制剂,OPN-6602具有克服MM的护理标准抗性机制的潜力,并且具有与其他治疗剂相结合的巨大潜力。

使用靶向和全基因组测序对 SCLC 患者的循环游离 DNA 进行分析
SCLC 肿瘤的基因组分析已发现大量拷贝数变异 (CNA) 和高突变率。 4、5 肿瘤抑制基因肿瘤蛋白 p53 基因( TP53 )和视网膜母细胞瘤 1 基因( RB1 )几乎普遍失活和普遍丢失,转录调节因子(SRY-box 转录因子 2 基因 [ SOX2 ]、核因子 IB 基因 [ NFIB ]、v-myc 禽类粒细胞瘤病毒致癌基因肺癌衍生同源基因 [ MYCL1 ]、v-myc 禽类粒细胞瘤病毒致癌基因神经母细胞瘤衍生同源基因 [ MYCN ] 和 v-myc 禽类粒细胞瘤病毒致癌基因同源基因 [ MYC ])频繁扩增,Notch 和组蛋白乙酰转移酶基因(CREB 结合蛋白基因 [ CREBBP ] 和 E1A 结合蛋白 p300 基因 [ EP300 ])反复突变。 4-6 肿瘤活检,尤其是连续活检,对于 SCLC 来说是一项重大挑战,7,8