XiaoMi-AI文件搜索系统
World File Search SystemCaspase

caspase 1抑制剂VX-765降低免疫激活,...
摘要HIV-1感染会导致炎性体的激活,这可能促进病毒传播并建立病毒储层。我们评估了caspase-1抑制剂VX-765对植入人类CD34 +造血干细胞的人源化NSG小鼠中HIV-1感染的影响。在HIV-1感染后的第1和第3天之间,淋巴结和骨髓中caspase-1,NLRP3和IL-1β的表达增加(平均折叠变化(FC)分别为2.08、3.23和6.05,p <0.001)。IFI16和AIM2表达在第24天达到峰值,并与IL-18水平升高(6.89 vs 83.19 pg/ml,p = 0.004),病毒载量和CD4 + T细胞的增加(分别为P <0.005和P <0.0001),分别为脾脏)。用VX-765处理第11天(0.47 vs 2.2 pg/ml,p = 0.045),第22天(7.8 vs 23.2 pg/ml,p = 0.04),显着降低了TNF-α(0.47 vs 2.2 pg/ml,p = 0.045),IL-18 p = 0.027)和脾脏中的总HIV-1 DNA(1 054 vs 2 889副本 /10 6个细胞,p = 0.029)。我们证明,感染后早期靶向炎性体激活可能代表了针对HIV治疗的治疗策略,以防止CD4 + T细胞耗竭并减少免疫激活,病毒载量和HIV-1储层形成。

肥胖或超重个体脑内 caspase 表达的免疫组织化学分析
Acta BBA ‐ Mol Basis Dis 。2017;1863(2):499-508。https://doi.org/10.1016/j.bbadis.2016.10.006 2. Hersey M、Woodruff J、Maxwell N 等人。高脂饮食会诱发神经炎症并降低肥胖大鼠海马对依他普仑的血清素反应。脑行为免疫。2021;96:63-72。https://doi.org/10.1016/j.bbi.2021.05.010 3. Wakabayashi T、Yamaguchi K、Matsui K 等人。饮食和基因诱导的大脑胰岛素抵抗对阿尔茨海默病小鼠模型中淀粉样蛋白病理的不同影响。 Mol Neurodegener。2019;14(1):15。https://doi.org/10.1186/s13024‐019‐0315-7 4. Zeyda M、Stulnig TM。脂肪组织巨噬细胞。Immunol Lett。2007;112(2):61-67。https://doi.org/10.1016/j.imlet.2007.07.003 5. Hahm JR、Jo MH、Ullah R、Kim MW、Kim MO。代谢应激改变抗氧化系统,抑制脂联素受体 1 并在小鼠脑中诱发类似阿尔茨海默氏症的病理。Cells。2020;9:249。 https://doi.org/10.3390/cells9010249 6. Mosser DM, Edwards JP。探索巨噬细胞活化的全部范围。Nat Rev Immunol。2008;8(12):958-969。https://doi.org/10. 1038/nri2448 7. Agustí A, García‐Pardo MP, López‐Almela I 等人。肠脑轴、肥胖和认知功能之间的相互作用。Front Neurosci。2018;12:155。https://doi.org/10.3389/fnins.2018.00155 8. Valdes AM, Walter J, Segal E, Spector TD。肠道菌群在营养和健康中的作用。BMJ。 2018;361:k2179。https://doi.org/10. 1136/bmj.k2179 9. Fricker M、Tolkovsky AM、Borutaite V、Coleman M、Brown GC。神经元细胞死亡。Physiol Rev。2018;98(2):813-880。https://doi. org/10.1152/physrev.00011.2017 10. Xu X、Lai Y、Hua ZC。细胞凋亡和凋亡小体:疾病信息和治疗靶点潜力。Biosci Rep。2019;39(1): BSR20180992。https://doi.org/10.1042/bsr20180992 11. Jan R、Chaudhry GE。了解针对细胞凋亡和凋亡途径的癌症治疗方法。Adv Pharm Bull。2019;9(2): 205-218。https://doi.org/10.15171/apb.2019.024 12. Green DR、Llambi F。细胞死亡信号。Cold Spring Harb Perspect Biol。2015;7(12):a006080。https://doi.org/10.1101/cshperspect.a0 06080 13. Khalifeh M、Penson PE、Banach M、Sahebkar A。他汀类药物作为抗焦亡药物。Arch Med Sci。2021;17(5):1414-1417。https://doi。 org/10.5114/aoms/141155 14. Winkler S、Rösen‐Wolff A。胱天蛋白酶-1:先天免疫的综合调节器。免疫病理学研讨会。2015;37(4):419-427。https://doi.org/ 10.1007/s00281-015-0494-4 15. Denes A、Lopez-Castejon G、Brough D。胱天蛋白酶-1:IL-1 只是冰山一角吗?细胞死亡研究。2012;3(7):e338。https://doi.org/10. 1038/cddis.2012.86 16. Makoni NJ、Nichols MR。胱天蛋白酶-1 活化的复杂生物物理谜题。生物化学与生物物理研究。 2021;15:108753。https://doi.org/ 10.1016/j.abb.2021.108753 17. Schmid‐Burgk JL、Gaidt MM、Schmidt T、Ebert TS、Bartok E、Hornung V。Caspase-4 介导人类髓系细胞中 NLRP3 炎症小体的非典型激活。Eur J Immunol。2015;45(10):2911-2917。 https://doi.org/10.1002/eji.201545523 18. Sankari SL、Masthan KM、Babu NA、Bhattacharjee T、Elumalai M。癌症中的细胞凋亡——更新。亚洲太平洋癌症预防杂志 APJCP 。 2012;13(10):4873-4878。 https://doi.org/10.7314/apjcp.2012.13.10。 4873 19. Gómez‐Apo E、Mondragón‐Maya A、Ferrari‐Díaz M、Silva‐Pereyra J. 与超重和肥胖相关的大脑结构变化。 J奥贝斯。 2021;2021:6613385-6613418。 https://doi.org/10.1155/2021/ 6613385 20. Herrmann MJ、Tesar A.-K、Beier J、Berg M、Warrings B. 肥胖中的灰质改变:全脑研究的荟萃分析。Obes Rev。2019;20(3):464-471。https://doi.org/10.1111/obr.12799
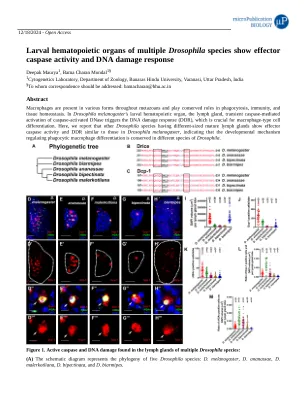
多种果蝇幼虫造血器官表现出效应胱天蛋白酶活性和DNA损伤反应
1882 年,埃利·梅契尼科夫 (Élie Metchnikoff) 在海星幼虫中发现了巨噬细胞,这种细胞通过吞噬外来物质来破坏外来物质。他将这一过程描述为吞噬作用 (Underhill 等人,2016)。后续研究表明,巨噬细胞在整个后生动物中都得到了保留,在调节发育、组织修复、体内平衡和先天免疫方面表现出额外的功能 (Lazarov 等人,2023;Park 等人,2022)。在三胚层动物中,吞噬细胞由于开放的循环系统而穿过体腔并清除细胞碎片或病原体 (Maheshwari,2022;Banerjee 等人,2019)。在哺乳动物中,常驻组织巨噬细胞在早期胚胎阶段从卵黄囊和红细胞-髓系前体细胞发育而来,并在整个生命过程中具有自我更新能力。单核细胞衍生的巨噬细胞也与快速补充的组织有关,例如肠道(Lazarov 等人,2023;Lee & Ginhoux,2022;Park 等人,2022)。在从单细胞生物进化到高度复杂的脊椎动物的过程中,巨噬细胞的作用和吞噬过程在很大程度上保持了下来(Yutin 等人,2009)。然而,吞噬巨噬细胞分化的潜在机制仍不清楚。

研究论文 kin17 缺乏通过调节 Caspase 3、PARP 和 Bcl-2 家族蛋白促进宫颈癌细胞凋亡
背景 .由于晚期宫颈癌的治疗手段不具特异性以及缺乏分子靶向药物,晚期宫颈癌的治疗仍具有较大的挑战性,寻找新的宫颈癌治疗生物标志物十分必要。方法 .本研究通过转染携带KIN17 siRNA的重组慢病毒载体,构建kin17敲低的宫颈细胞株HeLa和SiHa,并用嘌呤霉素进行筛选。通过荧光观察和蛋白质印迹法检测建立的kin17敲低细胞。流式细胞术检测细胞凋亡和线粒体膜电位(MMP)。分光光度法检测caspase 3酶活性。蛋白质印迹法分析凋亡相关蛋白的表达谱。最后,我们利用生物信息学和蛋白质组学数据分析宫颈癌中的KIN相关基因。结果 .结果显示,转染基因沉默载体的HeLa和SiHa细胞中kin17的荧光阳性率较高(> 90%),基因沉默效率较高(> 65%)。此外,kin17的缺失分别使HeLa和SiHa细胞的MMP降低和凋亡率增加。此外,敲低kin17可以增强HeLa和SiHa细胞中caspase 3酶活性,增加裂解PARP和Bim的表达,同时降低Bcl-xL和磷酸化BAD的表达。宫颈癌KIN相关预后基因的鉴定显示,共构建了5个基因(FZR1、IMPDH1、GPKOW、XPA和DDX39A)用于该风险评分,结果显示CTLA4表达与风险评分呈负相关。结论。我们的研究结果表明,kin17 敲低可通过靶向 caspase 3、PARP 和 Bcl-2 家族蛋白促进宫颈癌细胞凋亡。此外,kin17 可以通过线粒体途径调控癌细胞凋亡,可作为调节宫颈癌细胞凋亡的新型治疗靶点。

TIR 信号激活细菌中的 caspase 样免疫
胱天蛋白酶家族的蛋白酶以及 Toll/白细胞介素-1 受体 (TIR) 结构域蛋白在人类的先天免疫和调节细胞死亡中发挥着核心作用。在本研究中,我们描述了一种由胱天蛋白酶样蛋白酶和 TIR 结构域蛋白组成的细菌免疫系统。我们发现,一旦 TIR 蛋白识别出噬菌体入侵,它就会产生以前未知的免疫信号分子 ADP-环[N7:1′′]-核糖 (N7-cADPR)。这种分子特异性地激活细菌胱天蛋白酶样蛋白酶,然后无差别地降解细胞蛋白以阻止噬菌体复制。TIR-胱天蛋白酶防御系统(我们称之为 IV 型 Thoeris)在细菌中含量丰富,可有效防止噬菌体繁殖。我们的研究突出了 TIR 产生的免疫信号分子的多样性,并表明由胱天蛋白酶家族蛋白酶调节的细胞死亡是一种古老的先天免疫机制。

使用 RNAi 在中国仓鼠卵巢细胞中同时敲入/敲除 Caspase 8 相关蛋白 2 基因的时间和成本有效的基因组编辑方案
背景:CHO 细胞是生产生物制药的首选,而基因组编辑技术为提高重组蛋白产量提供了机会。靶向凋亡相关基因,如 Caspases 8 相关蛋白 2 (CASP8AP2),可提高 CHO 细胞的活力和生产力。将强大的策略与 CRISPR-Cas9 系统相结合使其能够应用于 CHO 细胞工程。目标:本研究旨在开发一种经济有效的方案,使用 CRISPR-Cas9 系统结合 HITI 策略同时在 CHO 细胞中缺失/插入 CASP8AP2 基因,并评估其对细胞活力和蛋白质表达的影响。材料和方法:我们通过将 CRISPR/Cas9 与 HITI 策略相结合,开发了一种有效的 CHO 细胞工程方案。使用 CHOPCHOP 软件设计了两个不同的 sgRNA 序列以靶向 CASP8AP2 基因的 3' UTR 区域。使用经济高效的 PEI 试剂将 gRNA 克隆到 PX459 和 PX460-1 载体中,并转染到 CHO 细胞中。采用手动选择系统简化单细胞克隆过程。MTT 测定评估 24、48 和 72 小时的基因沉默和细胞活力。流式细胞术评估 CASP8AP2 沉默的 CHO 细胞中的蛋白质表达。结果:研究证实了将 CRISPR-Cas9 与 HITI 策略相结合的稳健性,在产生敲除克隆方面实现了 60% 的高效率。PEI 转染成功地将构建体传递给近 65% 的克隆,其中大多数是纯合的。该方案被证明适用于资源有限的实验室,只需要倒置荧光显微镜。 CASP8AP2 敲除 (CHO-KO) 细胞经 NaBu 处理后,与 CHO-K1 细胞相比,其细胞存活率显著延长,48 小时时的 IC50 值分别为 7.28 mM 和 14.25 mM(P 值:24 小时 ≤ 0.0001,48 小时 ≤ 0.0001,P 值:72 小时 = 0.0007)。与天然细胞相比,CHO CASP8AP2 沉默细胞的 JRed 表达增加了 1.3 倍。结论:使用 CRISPR-Cas9 和 HITI 策略有效改造 CHO 细胞,同时进行 CASP8AP2 基因缺失/插入,从而提高细胞存活率和蛋白质表达。
线粒体动力学通过控制 caspase 依赖的 DNA 损伤来调节基因组稳定性
Kai Cao,1、2、3、9 Joel S. Riley,1、2、4、9 Rosalie Heilig,1、2 Alfredo E. Montes-Go´ mez,1、2 Esmee Vringer,1、2 Kevin Berthenet,5、6 Catherine Cloix,1、2 Yassmin Elmasry,1、2 David G. Spiller,7 Gabriel Ichim,5、6 Kirsteen J. Campbell,1、2 Andrew P. Gilmore,8 和 Stephen WG Tait 1、2、10、* 1 英国癌症研究中心 Beatson 研究所,英国格拉斯哥 G61 1BD 2 格拉斯哥大学医学、兽医学和生命科学学院癌症科学研究所,英国格拉斯哥 G61 1QH 3 北京工业大学环境与生命科学学院化学与生物系,北京 100124,中国 4 因斯布鲁克医科大学生物中心发育免疫学研究所,因斯布鲁克,奥地利 5 里昂癌症研究中心 (CRCL),INSERM 1052,CNRS 5286,里昂,法国 6 癌细胞死亡实验室,LabEx DEVweCAN 的一部分,里昂大学,法国里昂 7 系统显微镜,曼彻斯特大学生物、医学与健康学院,曼彻斯特 M13 9PT,英国 8 威康细胞基质研究中心,曼彻斯特学术科学中心,生物、医学与健康学院,曼彻斯特大学,曼彻斯特 M13 9PT,英国 9 这些作者贡献相同 10 主要联系人 * 通讯地址:stephen.tait@glasgow.ac.uk https://doi.org/10.1016/j.devcel.2022.03.019

与辐射和ATR抑制剂联合治疗后的免疫原性死亡受凋亡caspase双重调节
结果:相对于模拟治疗或单独使用辐射处理的细胞,在与辐射和ATR抑制剂联合处理后的72小时后,所有细胞系的细胞外释放均在所有细胞系中增加。HMGB1释放在很大程度上与质膜完整性的丧失相关,但并非严格相关,并通过添加caspase抑制剂而被抑制。然而,尽管caspase抑制了caspase,但在该细胞系caspase抑制诱导的PMLKL中,一条细胞系显示了HMGB1的释放,这是坏死性的标记。ATP分泌发生在共同治疗后的48小时内,显然与质膜完整性的丧失无关。添加pan-caspase抑制作用进一步增加了ATP分泌。在辐照后24-72小时时,钙网蛋白的表面呈递增加,但通过ATR或caspase抑制进一步增加。

高级制造的新欧洲计量网络
家族性阿尔茨海默氏症疾病(FAD)的常染色体显性形式是由淀粉样蛋白前光体蛋白(APP)基因的突变以及编码Presenilin 1或Presenilin 2的基因的突变引起的。同时,有证据表明,增加的氧化应激可能在瑞典时尚的快速发展中起着至关重要的作用。在这里,我们研究了 - 淀粉样蛋白前体蛋白对PC12细胞中氧化应激的细胞死亡机制的瑞典双突变(K670m/N671L)的影响。Western印迹分析和caspase底物的裂解研究在包含瑞典APP突变的细胞中用过氧化氢处理后,执行者caspase 3的活性升高。这种升高的活性是固有和外部凋亡途径的激活增强的结果,包括caspase 2和caspase 8的激活。此外,我们观察到通过保护线粒体功能障碍和降低caspase 9活性的JNK抑制剂SP600125(SP600125)对JNK途径的激活和凋亡的激活增强。我们的发现提供了证据,表明在幼年时代,大量的神经退行性因素可能是由于氧化应激水平升高而激活不同凋亡路径的脆弱性,因此神经元的脆弱性增加。

线粒体质量控制基因的破坏促进了凋亡刺激后caspase耐药细胞的存活
在经历细胞内凋亡的细胞中,线粒体外膜通透性(MOMP)通常标志着细胞死亡过程中不可逆的一步。然而,在某些情况下,被处理的细胞的亚群可以表现出一个余生的反应,称为“少数MOMP。”在这种现象中,尽管caspase激活水平较低,并且随后受到核酸内切酶caspase激活的DNase(DNA片段化因子亚基β>因此,这些细胞会经历DNA损伤,从而增加了肿瘤发生的可能性。但是,对少数MOMP响应知之甚少。发现影响单个细胞中MOMP反应的基因,我们构成了基于成像的表型siRNA筛选。我们鉴定了多个候选基因,其下调增加了单个细胞内MOMP的异质性,其中是与线粒体动力学和线粒体有关的基因,该基因参与了线粒体质量控制(MQC)系统。此外,为了测试功能性MQC对于降低少数MOMP的频率很重要的假设,我们开发了一种测量caspase参与细胞的固定生存的测定法。我们发现,在各种MQC基因中表现出的细胞确实很容易出现在成型后生存。我们的数据突出了蛋白质与线粒体动力学和线粒体中有关的重要作用,在防止凋亡失调和肿瘤发生中。