XiaoMi-AI文件搜索系统
World File Search SystemCs

CS/CS/HB 107 电动汽车和插电式汽车的影响...
佛罗里达州是全美注册电动汽车数量第二多的州。电动汽车和混合动力汽车的市场份额不断增加,以及内燃机汽车的燃油效率不断提高,预计将继续对全州和地方的燃油税收入产生不利影响,而燃油税是交通支出的资金来源。在未来 20 年内,佛罗里达州因电动汽车市场渗透率提高而造成的收入损失估计在 5.6% 到 20% 之间,具体取决于电动汽车市场的渗透率。在地方层面,收入损失可能会影响当地道路和公共交通的运营和维护。目前,佛罗里达州的普通司机每年为交通相关项目缴纳 283 美元的机动车燃油税。佛罗里达州目前不对电动汽车或插电式混合动力汽车征收额外的注册费、消费税或使用费。该法案要求收入估算会议 (REC) 估算 2024-2025、2025-2026 和 2026-2027 财年对用于给电动汽车充电的电力销售征收的销售税对一般收入基金的影响。REC 必须在适用财政年度开始前的 6 月 10 日之前向税务部提供此估算。此规定将于 2027 年 6 月 30 日到期。从 2024 年 7 月开始,该法案指示税务部向州交通信托基金 (STTF) 分配 REC 为该州财政年度估算的金额的十二分之一。此规定将于 2027 年 6 月 30 日到期。此外,该法案认识到,电动汽车和插电式混合动力汽车的持续普及将对州税收和 STTF 产生重大影响。为了更好地理解这些影响,该法案要求经济和人口研究办公室 (EDR) 制作一份报告,估计对州交通信托基金的长期影响、预计继续在该州采用电动汽车和插电式混合动力汽车将对销售税和总收入税收产生的影响,以及使用住宅充电站而非商业充电站的用户百分比。该报告还必须考虑激励或推广电动汽车的联邦政策以及这些政策对电动汽车持续采用速度的影响。EDR 必须在 2026 年 12 月 1 日之前提交报告,2027 年立法机关必须使用该报告来考虑解决对 STTF 的长期影响所需的潜在政策变化。收入估算会议尚未估计该法案的收入影响。对于 2024-2025、2025-2026 和 2026-2027 财政年度,该法案将对一般收入产生负面影响,并对州信托基金收入产生同等的积极影响。

CS/CS/HB 619专业公司的主权免疫
摘要分析CS/CS/HB 619于2024年2月22日通过了房屋,随后于2024年3月4日通过了参议院。“主权豁免权”是一种学说,源自英国普通法,该法律禁止诉讼反对政府不在政府同意被起诉的情况下。此一般原则的例外包括自愿放弃主权豁免权,而当实体不是“国家的部门”时,也就是说,当主权免疫不适用时,因为从技术上讲,这不是国家被起诉的国家。除非适用,否则主权免疫学说违反了州或联邦法院针对佛罗里达州的诉讼,除非适用。但是,第X条,《佛罗里达州宪法》第13条授权立法机关放弃主权豁免权,“关于现在或以后的所有负债”。根据这项权力委派,立法机关在多个领域放弃了主权豁免权,包括在一定程度上进行指定的侵权诉讼。 768.28(1),F.S。佛罗里达州的法律还通过将这些侵权诉讼的国家主权豁免权扩展到某些私人实体,将其指定为“国家的代理人”。例如,s。 768.28(10)(e),F.S.,指定出于s的目的。 768.28(1),F.S.是一家专业公司,提供指定的监视和检查服务或执行此类服务的任何公司的任何雇员,是佛罗里达州交通部(“ FDOT”)的代理人,同时在公司合同范围内行动。法案修改。 768.28(10)(e),F.S。,以扩大授予专业公司及其雇员的主权豁免保护的范围。具体来说,该法案规定,主权豁免保护不仅适用于与FDOT直接合同的专业公司,还适用于提供与FDOT直接合同的专业公司的监控和检查服务的专业公司。此外,根据法案,与专业公司的任何合同都必须赔偿FDOT,包括任何责任,包括合理的律师费,直到s中规定的法定恢复限制。 768.28(5)(a),F.S.,在公司或其雇员的疏忽范围内。

CS 730/830
除了课本、讲座和背诵之外,您还必须完成作业。不允许使用 AI 编码助手(例如 GitHub CoPilot)(就像在编码面试中不允许使用一样)。如果您查阅网站或与他人深入讨论作业的概念,则应在作业中列出它们。不要与他人谈论您的作业代码。如果您作弊,您将获得不及格的课程成绩,并被报告给系主任和 CEPS 院长。如果您发现情绪或心理健康问题影响了您的表现,请随时联系咨询中心 www.unh.edu/pacs — 他们通常会有所帮助!时间管理技能的资源是 www.unh.edu/cfar。如果宗教信仰会妨碍您按时完成作业,请在学期开始时告知我,以便我提供适当的安排。如果您已注册学生无障碍服务,请让他们给我发一封信,并请尽快与我交谈,以便我提供适当的安排。如果您因病需要隔离,请尽快告知我您预计何时可以返回课堂。

CS网站PL Michael J. Napuda,Esq。 114222014 Masten和Ray 254 South Broadway; P.O. Box 406 Pennsville,NJ 08070电话(856)678-4777;传真(856)678 6805律师 议程计划委员会 健康的Maywood 的元素
18 Valley Grove商业园区平台Q 1234 S Valley Grove Way细分Plat The Grove -Changange正在审查,最新提交11/1/24



计算机科学 (CS)
CS 1428。计算机科学基础 I。面向计算机科学专业、辅修专业和其他希望了解计算机科学技术入门的学生的入门课程。包含数字计算机的历史和结构概述,包括二进制数据表示。强调问题解决、算法开发、结构化编程、良好的编码风格和 C++ 的控制结构。先决条件:[MATH 1315 或 MATH 1317 或 MATH 1319 或 MATH 1329 或 MATH 2417 或 MATH 2471,成绩为“C”或更高] 或 [ACT 数学成绩为 24 或更高] 或 [新 ACT 数学成绩为 25 或更高] 或 [SAT 数学成绩为 520 或更高] 或 [SAT 数学部分成绩为 550 或更高] 或 [Accuplacer College 数学成绩为 86 或更高] 或 [Compass College 代数成绩为 46 或更高] 或 [下一代高级代数和函数测试为 263 或更高]。4 个学分。3 个讲座接触时间。2 个实验室接触时间。课程属性:Dif Tui- 科学与工程|实验室要求成绩模式:标准字母 TCCN:COSC 1437

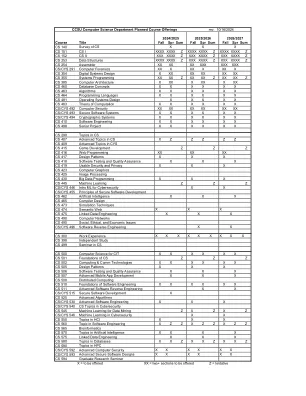
课程标题CS 140 CS X X X CS 151 CS I XXXX XXXX Z XXXX Z XXXX XXXX Z XXXX XXXX XXXX Z CS 152 CS 152 CS II XXXX XXXX ZXXX Z XXXX XXXX XXXX ZXXX ZXXX XXX Z xxxx Z CS CS CS CS 253数据Struc
课程标题CS 140 CS X X X CS 151 CS I XXXX XXXX Z XXXX XXXX XXXX Z XXXX XXXX XXXX Z CS CS 152 CS II XXXX XXXX ZXXX ZXXX XXXX XXXX XXXX ZXXX Z CS CS CS CS CS CS CS CS CS 253数据结构 Assembler XX XX XX XXX XXX XXX CS/CYS 291 Computer Forensics XX X XX X XX X CS 354 Digital Systems Design X XX XX XX XX XX CS 355 Systems Programming XX XX Z XX XX Z XX XX Z CS 385 Computer Architecture X XX X XX X XX CS 460 Database Concepts X X X X X X CS 463 Algorithms X X X X X X CS 464 Programming Languages X X X X X X CS 481 Operating Systems Design X X X X X CS 483 Theory of Computation X X X X X X CS/CYS 492 Computer Security XX XX XX XX XX XX CS/CYS 493 Secure Software Systems X X X X X X CS/CYS 494 Cryptographic Systems X X X X X X CS 410软件工程X X X X X X CS 498高级项目X X X X X X X