XiaoMi-AI文件搜索系统
World File Search SystemDHFR

通过抑制DHFR和TRXR在乳腺癌细胞中的DHFR和TRXR的多靶标抗癌活性
评估了由硫烷金(I)化合物(I)化合物,由偶氮和磷酸盐配体制成,以筛查乳腺癌细胞板(BC)的筛查。这些化合物在中央金属周围具有N-AU-P或CL-AU-P键,并且在唑拓和/或磷酸基部分中存在质子或原始极性基团以调整其亲水性。在六个候选物中,只有具有P-AU-N环境的化合物,并且未发现配体中的羟基和羧基的化合物。The compounds were screened by MTT tests in SKBR3, A17, and MDA-MB231 cancer cells, and two compounds (namely the 4,5-dicyano-imidazolate-1yl-gold(I)-(triphenylphosphane, 5, and 4,5-dichloro-imidazolate-1yl-gold(I)-triphenylphosphane, 6) were found very细胞毒性,在MDA-MB231细胞中最活跃的50值为3.46 µm通过在处理的细胞裂解物中进行酶促测定,与对照细胞相比,已经在细胞处理后4或12小时测量了二氢叶酸还原酶(DHFR)的残留酶活性。在处理12小时后,SKBR3和A17细胞的DHFR活性显着降低,而化合物5和6,但在人类MDA-MB231细胞中却没有显着降低。有趣的是,在治疗4小时后发现它非常高,揭示了DHFR酶试验的时间依赖性。DHFR抑制数据已与硫氧还蛋白还原酶(TRXR)的DHFR抑制数据(TRXR)是金化合物的最公认的分子靶标。对于后者,发现了类似的残留活性(即SKBR3细胞和化合物5或6的匹配分别为37%和49%)。对CT-DNA(CALF-thymus-DNA)和血浆转运蛋白(例如BSA(牛血清白蛋白)和ATF(APO转移蛋白))的结合研究进行了研究。对金化合物的预期,数据支持与蛋白质的强结合(K SV值范围:1.51÷2.46×10 4 m-1),与CT-DNA的次要凹槽(K SV值范围:1.55÷6.12×10 3 M-1)的相互作用较弱。

结核分枝杆菌二氢叶酸还原酶抑制剂:过去20年的先进状态
摘要:结核病是一种具有高发病率和死亡率的疾病。由于与当前疗法有关的问题,开发新药进行治疗是必要和迫切的。二氢叶酸还原酶(DHFR)是多种药物作用的公认靶标。与人DHFR(H DHFR)相比,结核分枝杆菌DHFR(MT DHFR)DHFR(MT DHFR)的3D结构阐明了配体特异性的主要氨基酸残基和结构基础。本文旨在就过去二十年来开发的新MT DHFR抑制剂提供有关最先进的观点。这项研究表明,功效与特定组的存在之间的相关性,例如与酶活性位点结合的二氨基吡啶环与经典DHFR抑制剂甲氨蝶呤的结合的相似性。在此,还报道了最近开发分子非传统核心的努力,这可能更有选择性和有效地抵抗结核病。

与诊断和发展阿尔茨海默氏病有关的Circ_0001535的原始研究生物信息学和实验鉴定
背景:阿尔茨海默氏病(AD)是一种疾病,经常发生在老年人群中。仍然缺乏针对这种疾病的诊断和治疗方法,需要进行更多的研究。此外,对AD中的圆形RNA(CIRCRNA)的功能知之甚少。方法:在这项研究中,下载了基因表达综合(GEO)数据库的AD的RNA表达数据。通过逆转录质量PCR(RT-QPCR)测量健康参与者和AD患者脑脊液样品中CIRCRNA的表达水平。用接收器操作特征曲线(ROC)分析了差异表达的CIRCRNA的诊断值。途径。使用染色质免疫沉淀(CHIP)和RNA结合蛋白免疫沉淀(RIP)分析,验证了Circ_0001535和E2F转录因子1(E2F1)或E2F1和二氢叶酸还原酶(DHFR)之间的直接相互作用。细胞计数KIT-8(CCK8)和流式细胞仪用于识别CIRC_0001535/E2F1/DHFR轴对AD细胞增殖和凋亡的功能。结果:总共有12个CIRCRNA与AD诊断有关。AD患者和对照组之间7个CIRCRNA的表达水平有所不同。CRIC_0001535在这些CIRCRNA中具有最大的诊断价值。因此,在本研究中,Circ_0001535被视为关键的Circrna。E2F1/DHFR轴预计由Circ_0001535调节。E2F1/DHFR轴预计由Circ_0001535调节。此外,IP分析实验结果表明,E2F1可以与DHFR的启动子区域结合,并受CIRC_0001535的调节。体外结果表明,Circ_0001535过表达可以促进DHFR的表达,而E2F1敲低可以抑制SH-SY5Y细胞中的DHFR表达。最后,救援实验表明,Circ_0001535可以减少β25-35诱导的SH-SY5Y细胞增殖,并通过E2F1/DHFR轴促进凋亡。结论:我们在广告中的研究可以提供有关特定ciRCRNA在AD环境中的作用的重要信息,并指出AD中治疗干预的特定未来领域。

药物名称:甲氨蝶呤
甲氨蝶呤是一种叶酸拮抗剂。2 四氢叶酸是叶酸的活性形式,是嘌呤和胸苷酸合成所必需的。叶酸被二氢叶酸还原酶 (DHFR) 还原为四氢叶酸。甲氨蝶呤的细胞毒性来自三种作用:抑制 DHFR、抑制胸苷酸和改变还原叶酸的转运。3 抑制 DHFR 会导致胸苷酸和嘌呤缺乏,从而导致 DNA 合成、修复和细胞复制减少。3 DHFR 对甲氨蝶呤的亲和力远大于其对叶酸或二氢叶酸的亲和力,因此同时给予大剂量叶酸不会逆转甲氨蝶呤的作用。 2 然而,如果在甲氨蝶呤后不久服用四氢叶酸衍生物亚叶酸钙,则可能会阻断甲氨蝶呤的作用,因为它不需要 DHFR 来激活。2 中等剂量 (> 100 mg/m 2 ) 至高剂量甲氨蝶呤 (> 1000 mg/m 2 )4 加亚叶酸救援通常用于癌症治疗。3 甲氨蝶呤对快速增殖细胞最有效,因为细胞毒作用主要发生在细胞周期的 S 期。3 甲氨蝶呤还具有免疫抑制活性,可能是由于抑制淋巴细胞增殖。5

二氢叶酸还原酶活性控制发育中的新皮质中的神经源性转变
一碳/叶酸 (1C) 代谢提供 DNA 和组蛋白甲基化所需的甲基基团,并参与维持干细胞的自我更新。二氢叶酸还原酶 (DHFR) 是 1C 代谢中的关键酶,在新皮层发育的早期阶段,在人类和小鼠神经祖细胞中高度表达。在这里,我们研究了 DHFR 在发育中的新皮层中的作用,并报告说,降低其在人类神经类器官和小鼠胚胎新皮层中的活性会加速间接神经发生,从而影响新皮层的神经元组成。此外,我们表明,降低神经祖细胞中的 DHFR 活性会导致一碳/叶酸代谢物的减少,并与 H3K4me3 水平的变化相关。我们的研究结果揭示了 DHFR 在控制新皮层发育的特定步骤中发挥的意想不到的作用,并表明 1C 代谢线索的变化会影响细胞命运转变。

主动脉瘤
摘要:我们已经证明,内皮特异性 DHFR(二氢叶酸还原酶)缺乏是 eNOS(内皮 NO 合酶)解偶联和腹主动脉瘤 (AAA) 形成的原因。在本文中,我们研究了 microRNA-192-5p 在介导 NOX(NADPH 氧化酶)依赖性 DHFR 缺乏和 AAA 形成中的新作用。microRNA-192-5p 预计以 DHFR 为靶点。有趣的是,人类 AAA 患者的智人 - microRNA-192-5p 表达显著上调。在暴露于过氧化氢 (H 2 O 2 ) 的人主动脉内皮细胞中,智人 - microRNA-192-5p 表达显著上调。这伴随着 DHFR mRNA 和蛋白质表达的显著下调,而智人 - microRNA-192-5p 特异性抑制剂可恢复这种下调。值得注意的是,microRNA-192-5p 表达在血管紧张素 II(血管紧张素 II)输注的 hph-1(高苯丙氨酸血症 1)小鼠中显著上调,而在 hph-1–NOX1、hph-1–NOX2、hph-1–中性粒细胞胞质因子 1 和 hph-1–NOX4 双突变小鼠中减弱,AAA 发病率也消失,表明 microRNA-192-5p 在 NOX 激活后具有下游效应作用。在超声和尸检中,用小鼠–microRNA-192-5p 抑制剂进行体内治疗可减弱血管紧张素 II 输注的 hph-1 小鼠的腹主动脉扩张。它还逆转了血管重塑的特征,包括基质降解、外膜肥大和腔内血栓形成。这些动物恢复了 DHFR mRNA 和蛋白质表达,减弱了超氧化物的产生,重新偶联了 eNOS,并保留了 NO 的生物利用度。总之,我们的数据首次证明了 microRNA-192-5p 在介导 NOX 依赖性 DHFR 缺乏和 AAA 形成中起着关键作用,抑制 DHFR 缺乏和 AAA 形成可有效减缓 AAA 的发展。由于小鼠和人类 microRNA-192-5p 序列相同,microRNA-192-5p 抑制剂可能很容易转化为治疗 AAA 的新型疗法。(高血压。2021;78:282–293。DOI:10.1161/HYPERTENSIONAHA.120.15070。)• 数据补充

与低剂量甲氨蝶呤的astaxanthin共同治疗增加了细胞周期停滞,并改善NALM-6
甲氨蝶呤(MTX),广泛识别的化学疗法药物,通过抑制包括叶酸途径中的各种酶(包括二氢叶酸还原酶(DHFR))的各种酶来抑制DNA合成。MTX被认为是所有不同治疗方案中的关键元素(4)。DHFR产生THF,而胸甲酯合成酶(TYMS)则利用亚甲基THF作为其底物。Tyms在DNA合成和修复中起重要作用(5)。Tyms通过抑制甲氨蝶呤的作用来促进抗增生特性(6);因此,已利用TYMS水平预测MTX治疗结果(7)。尽管甲氨蝶呤在所有人的治疗中都非常有效,但它有几个缺点,其中一些可能是威胁生命的(8,9)。进行化学疗法作为白血病治疗的常规方法与许多局限性有关。此外,MTX耐药性为所有化学疗法的成功带来了重要的障碍(8-10)。

用于特定细胞靶向和药物输送的多功能淀粉样蛋白寡聚纳米粒子
116 试剂和酶。除非另有说明,试剂和酶均从 Sigma-Aldrich(英国)购买。碳网格(400 平方目铜)从 Micro to Nano(荷兰)购买,醋酸铀酰溶液由巴塞罗那自治大学的显微镜服务部门提供。Sup35- 121 SAC 肽从 CASLO ApS(Scion 丹麦技术大学)购买。122 蛋白质的表达和纯化。克隆到带有 His6 标签的质粒 pET28(a) 中的 Sup35- 123 5aa-DHFR 的 cDNA 是从 GenScript 获得的。通过在 128 质粒 pET28(a)/Sup35-5aa-DHFR 上进行诱变,获得了构建体 pET28(a)/Sup35-8aa- 126 DHFR、pET28(a)/野生型 DHFR (DHFR-wt) 和 pET28(a)/ 127 Sup35-5aa-DHFR-Z。用相应的质粒转化大肠杆菌 BL21 (DE3)- 129 感受态细胞。130 然后,将转化细胞在 10 mL 溶源性肉汤 (LB) 中培养
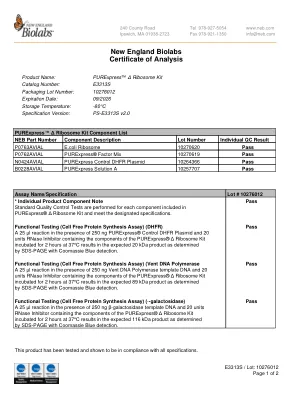
新英格兰生物实验室分析证书
在 250 ng PURExpress® 对照 DHFR 质粒和 20 单位 RNase 抑制剂(含有 PURExpress® Δ 核糖体试剂盒的成分)存在下进行 25 µl 反应,在 37°C 下孵育 2 小时,通过 SDS-PAGE 和考马斯亮蓝检测测定,可得到预期的 20 kDa 产物。
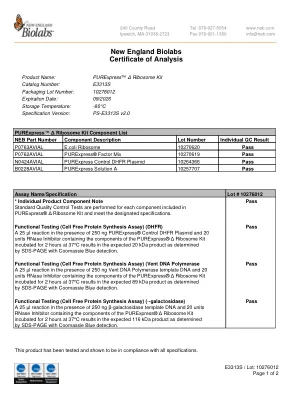
新英格兰Biolabs分析证书
A 25 µL反应和20个单位RNase抑制剂,其中含有Purexpress®Δ核糖体的成分在37°C下在37°C下孵育2小时,从而在预期的20 kDa产物中通过SDS-PAGE与Coomassie Blue blue detection确定,导致预期的20 kDa产物。