XiaoMi-AI文件搜索系统
World File Search SystemDNMT3A

癌症治疗:表观遗传学观点 - Thieme Connect
正常细胞中的 DNA 甲基化和组蛋白修饰 DNA 甲基化是最著名、研究最全面的表观遗传机制。DNA 甲基化的主要作用是阻止基因表达。DNA 甲基化意味着在胞嘧啶核苷酸的 5′ 位置共价添加一个甲基 (-CH 3 )。1,2 负责添加甲基的酶称为 DNA 甲基转移酶 (DNMT)。哺乳动物有五种 DNMT,DNMT1、DNMT2、DNMT3a、DNMT3b 和 DNMT3L。其中,只有 DNMT1、DNMT3a 和 DNMT3b 可以将甲基从 S-腺苷甲硫氨酸 (SAM) 转移到 DNA 上。3 DNMT1 负责维持 DNA 甲基化。在复制过程中,DNMT1 转录新合成链上预先存在的甲基化标记。3 然而,在体内研究中,DNMT1 已被证明

单细胞 DNA 测序揭示了移植合格多发性骨髓瘤患者中 CHIP 的进化模式
平台,具有 20 个白血病相关基因的目标面板。我们在移植时检测到 6/12 名患者 (50%) 的 CHIP 致病突变。最常发生突变的基因是 TET2 、 EZH2 、 KIT 、 DNMT3A 和 ASXL1 。在两名患者中,我们观察到同一克隆中同时发生涉及表观遗传修饰因子 (即 DNMT3A) 和/或涉及剪接机制的基因 (即 SF3B1) 和/或酪氨酸激酶受体 (即 KIT) 的突变。配对样本的纵向分析显示,随着时间的推移,突变的高适应度克隆会得到正向选择,无论它们与主要或次要亚克隆的亲和力如何。所有基因的拷贝数分析未显示干细胞区室中存在任何数值改变。此外,我们观察到 CHIP 阳性患者与未阳性患者相比,对治疗的反应往往不太理想。证实了高适应度突变随时间的亚克隆动态。

靶向染色质编辑揭示的模拟表观遗传记忆
图 2. DNMT3A 编辑细胞中的基因表达动态表明了一种不同于二进制的记忆形式。A 使用与 dCas9、PhlF 或 rTetR 融合的 KRAB、DNMT3A 或 TET1 作为 DNA 结合域 (DBD) 进行瞬时表观遗传编辑的概述。B 本研究开发的实验系统示意图。报告基因通过位点特异性染色体整合整合到内源性哺乳动物基因座中。哺乳动物组成型启动子 (EF1a) 驱动荧光蛋白 EBFP2 的表达。上游结合位点能够靶向募集表观遗传效应物,这些效应物与 DNA 结合蛋白 rTetR、PhlF 或 dCas9 融合。报告基因两侧是染色质绝缘体,以与其他基因隔离。 C 实验概述描述了瞬时转染到带有报告基因的细胞、基于转染水平的荧光激活细胞分选和时间过程流式细胞术测量。D 根据图 C 中显示的实验时间线,DNMT3A 编辑(DNMT3A-dCas9)报告基因的基因表达动态。显示的是 DNMT3A 编辑细胞的单细胞流式细胞术测量(EBFP2)。DNMT3A-dCas9 靶向启动子上游的 5 个靶位点,并使用乱序 gRNA 靶序列作为对照(图 SE.2 A、B、表 S3)。黄色阴影表示检测到转染标记的时间。显示的数据来自 3 个独立重复的代表性重复。E 转染 DNMT3A-dCas9 和细胞分选后 14 天进行 MeDIP-qPCR 和 ChIP-qPCR 分析,以获得高水平的转染。分析了启动子区域(表 S4 和方法)。显示的数据来自三个独立的重复。报告的是使用标准 ∆∆ C t 方法相对于活性状态的倍数变化及其平均值。误差线是平均值的标准差。DNMT3A-dCas9 靶向启动子 (gRNA) 上游的 5 个靶位点。使用乱序的 gRNA 靶序列 (gRNA NT) 作为对照。* P <0.05,** P <0.01,*** P <0.001,非配对双尾 t 检验。F 根据图 C 中显示的实验时间线的 KRAB 编辑 (PhlF-KRAB) 基因表达动态。显示的是单个细胞的报告基因 (EBFP2) 的流式细胞术测量值。黄色阴影区域表示在未应用 DAPG 期间检测到转染标记的时间。从第 6 天开始,在 PhlF-KRAB 和 PhlF 条件下应用 DAPG。每天测量不同的独立重复。显示的数据来自 3 个独立重复。G 转染 PhlF-KRAB 和高水平转染细胞分选后 6 天的 MeDIP-qPCR 和 ChIP-qPCR 分析。分析的是启动子区域。数据来自三个独立重复。显示的是相对于活性状态的标准 ∆∆ C t 方法确定的倍数变化及其平均值。误差线是平均值的标准差。* P <0.05,** P <0.01,*** P <0.001,非配对双尾 t 检验。H 当 KRAB = 0、TET1 = 0 时获得的染色质修饰回路。参见 SI 图 SM.1 C。I 上图:(CpGme, X) 对的剂量反应曲线。下图:DNMT3A 脉冲强度与 DNA 甲基化等级 (CpGme) 之间的剂量反应曲线。脉冲强度通过增加其高度来增加。参见 SI 图 SM.1 D 和 SM.3。J 系统基因表达的平稳概率分布,由 SI 表 SM.1 和 SM.4 中列出的反应表示,参数值在 SI 第 S.9.3 节中给出。K 系统在 t = 28 天后的基因表达概率分布,如图 J 所示,参数值和初始条件在 SI 第 S.9.4 节中给出。参见 SI 图 SM.1 B 和 SM.2。在图 I 和 J 中,DNMT3A 动力学被建模为随时间呈指数下降的脉冲(参见第 S.1.1 节 - SI 方程 (SM.7))。在我们的模型中,ε (ζ) 是衡量基础(招募)擦除率与每次修饰的自催化率之间比率的参数。参见 SI 图 SM.1 E 和 SM.3。

通过CRISPR/CAS9
为了研究克隆造血性基因突变的体外基因突变,并揭示了对人类茎和祖细胞(HSPC)室的直接影响,我们针对健康的,年轻的造血祖细胞,该细胞源自脐带血液样本,并用CRASPR/CAS9技术来源。位点特异性突变,随后在短期和长期的体外培养试验中分析,以评估自我更新和差异能力。菌落形成单元(CFU)测定法显示,TET2突变(TET2 MUT)细胞的自我更新增强,而ASXL1 MUT以及DNMT3A MUT细胞的自我更新并未揭示短期培养的显着变化。引人注目的是,在所有突变体的长期培养实验中都可以检测到增强的菌落形成,表明自我更新能力的增加。尽管我们还可以证明所有突变体的不同细胞克隆的优先克隆膨胀,但长期培养后的克隆组成揭示了对HSPC的突变特异性影响。因此,通过使用原发性脐带血细胞,我们能够研究表观遗传驱动器突变,而不会混淆年龄或复杂的突变景观,而我们的发现为克隆血肿相关突变对人类茎和前代细胞的自治和核心组成的直接影响提供了证据。

引用已发表版本(APA):Hellesøy,M.,Engen,C.,Grob,T.,Löwenberg,B.,Valk,P.J.M。,&Gjertsen,B。T.(2021)。
发病率,急性髓样白血病(AML)的分子表现和结果受到性的影响,但很少关注男性和男性患者之间与性别相关的分子和表型差异无关。虽然发病率增加和风险较差与男性表型相关,但预后的FLT3内部串联复制(FLT3-ITD)突变差,并且与NPM1和DNMT3A的共突变在女性AML中的表现过高。在这里,我们已经通过临床数据,突变程序,基因表达和离体药物敏感性来投资性别与FLT3-ITD突变状态之间的关系:BEAT AML,LAML-TCGA和两个独立的Hovon/Sakk群体,包括1755 Aml Aml患者。我们发现普遍的性别相关分子差异。flt3 -ITD,NPM1和DNMT3A突变的共发生在女性中的代表性过高,而具有FLT3 -ITD的雄性的特征是RNA剪接和表观遗传模拟器基因中的其他突变。我们观察到多种白细胞相关基因以及伴随性的离体药物反应的分歧表达,暗示了离散功能特性。重要的是,仅在女性FLT3 -ITD -ITD -Mutated AML中观察到显着的预后。因此,我们建议以性调整的方式优化FLT3-ITD突变状态作为临床工具,并假设可以通过包括性别特定的考虑因素来改善AML治疗策略的预测,预测和发展。

通过分级DNA甲基化的模拟表观遗传细胞记忆
图2。DNMT3A募集后的基因表达动力学与数字记忆不一致。使用特定于特定于染色体的染色体整合的169个报告基因基因的示意图。哺乳动物170构成启动子(EF1A)驱动荧光蛋白EBFP2的表达。上游结合位点可实现靶向171的表观遗传效应子,该效应子与DNA结合蛋白RTETR融合在一起,PHLF或DCAS9。报告基因是由染色质绝缘子与其他基因分离出来的172。b实验概述,描述了瞬时转染到具有报告基因的173个细胞,基于转染水平的荧光激活的细胞分选,以及时间顺序的流量细胞仪174测量。根据面板中所示的175个实验时间表。显示的是四种不同水平的转染水平的报告基因176(EBFP2)的流量细胞仪测量值的分布。DNMT3A-DCAS9靶向启动子上游的5个目标位点,177用作炒GRNA目标序列作为对照(图se.2 a,b,表S3)。显示的数据来自来自3个独立重复的代表性178重复。d使用DNMT3A-179的流量细胞仪的单细胞基因表达测量值对应于面板C中所示的细胞(30天)。父母是指带有180个报告基因的未转染细胞。数据来自3个独立重复的代表性重复。平均值。e MedIP-QPCR和ChIP-QPCR 181分析DNMT3A-DCAS9和细胞分类后14天分析高水平的转染。分析了启动子区域182。显示的数据来自三个独立的重复。报道的是折叠变化及其平均值,使用183标准∆ ∆ c t方法相对于活性状态。错误条为S.D.DNMT3A-DCAS9的靶向位置为184至5个目标位点(GRNA)。使用炒GRNA目标序列(GRNA NT)作为对照。185 *p≤0.05,**p≤0.01,***p≤0.001,未配对的两尾t检验。根据面板中所示的实验时间线,krab抑制的基因表达动力学(PHLF-KRAB)186。所示是从四种不同水平的转染水平的187个报告基因基因(EBFP2)的流量细胞仪测量值的分布。每天测量一个独立的重复。显示的数据188来自3个独立重复。g重新激活细胞的百分比(400-10 5基因表达A.U.F.)对应于F. h Medip-QPCR面板中显示的189个细胞种群和CHIP-QPCR分析后6天对PHLF-KRAB和Cell 190排序进行了高水平的转染。分析是启动子区域的。数据来自三个独立的重复。191显示的是折叠变化,其平均值由标准∆ΔCT方法确定相对于活性状态。错误192条是S.D.平均值。p≤0.05,**p≤0.01,***p≤0.001,未配对的两尾t检验。参见SI图参见Si无花果。202i简化染色质修饰193当krab = 0,dnmt3a = 0,tet1 = 0时获得的电路图,而H3K9me3并未介导从头催化194 DNA甲基化的催化。SM.1 C. J顶图:(CPGME,H3K4ME3)对的剂量响应曲线。底部图:(DNMT3A,CPGME)对的剂量-195响应曲线。SM.1 D和SM.3。 k k的基因表达的概率分布196的系统,该系统由Si Tape Sm.1和Sm.3中列出的反应表示。 参见Si无花果。 SM.1 B和SM.2。 l概率197在t = 28天后的基因表达分布,如面板I所述获得。 在小组j和l中,将198 DNMT3A动力学建模为脉冲,随着时间的流逝会呈指数减小。 在我们的模型中,α'是通过抑制组蛋白修饰的DNA甲基化建立的归一化速率199,DNA甲基化擦除率200速率与激活组蛋白的擦除速率和激活的组蛋白修改速率之间的µ'是每个基准级别(ε')的级别(均为基础率(均))(招募)(招募)(招募)。修改。 参见SI图 SM.1 E和SM.3。SM.1 D和SM.3。k k的基因表达的概率分布196的系统,该系统由Si Tape Sm.1和Sm.3中列出的反应表示。参见Si无花果。SM.1 B和SM.2。 l概率197在t = 28天后的基因表达分布,如面板I所述获得。SM.1 B和SM.2。l概率197在t = 28天后的基因表达分布,如面板I所述获得。在小组j和l中,将198 DNMT3A动力学建模为脉冲,随着时间的流逝会呈指数减小。在我们的模型中,α'是通过抑制组蛋白修饰的DNA甲基化建立的归一化速率199,DNA甲基化擦除率200速率与激活组蛋白的擦除速率和激活的组蛋白修改速率之间的µ'是每个基准级别(ε')的级别(均为基础率(均))(招募)(招募)(招募)。修改。参见SI图SM.1 E和SM.3。SM.1 E和SM.3。
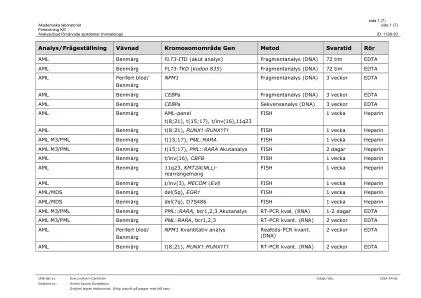
分析提供获得的疾病(血液学)
leukemiutringbenmärg/blod mylood面板*(ABL1,ANKRD26,ASXL1,ATRX,BCOR,BCOR,BCORL1,BRAF,CALR,CALR,CBL,CBL,CBL,CBL,CDKN2A,CDKN2A,CEBPA,CEBPA,CEBPA,CSF3R,CSF3R,CSF3R,CUX1,DDX41,DDX41,DNMT3A fbxw7, FLT3, GATA1, GATA2, GNAS, HRAS, Idh1, Idh2, Ikzf1, jak2, jak3, kdm6a, kit, kraas, kmt2a, mpl, myd88, NF1, Notch1 (INKLUSIVE 3´UTR), NPM1, NRAS, PDGFRA, PHF6, PPM1D, Pten, Ptpn11, Rad21, Runx1, Samd9, SAMDL9, Setbp1, SF3B1, SMC1A, SMC3, SRSF2, Stag2, Stat3, Stat5B, Tet2, TP53, U2AF1, WT1, ZRSR2, BTK, plcg2, terc) Div>

5月21日,星期二
p29。Isabelle Becker Megakaryocytes通过Rhoa Boston儿童医院和哈佛医学院P30的下游分泌自噬释放TGFβ1。Joyeeta Chakraborty化学基因植物,以定义Runx介导的转录调节电路Albert Einstein医学院P31。SETBP1中的Samantha Tauchmann突变增加了粒细胞谱系的输出,并激活与增殖相关基因骑士癌症研究所p32的转录。estelle carminita骨髓重塑和促炎性巨核细胞在波士顿儿童医院P33的慢性肾脏病鼠模型中。Nadia Carlesso上调造血干细胞中应力反应途径和镰状细胞疾病中的骨髓生态位。贝克曼研究所P34。sanika gupte中性粒细胞衍生的Sema4a是一种非细胞自动构成的骨髓骨膜自动调节剂,可保留髓样偏置的HSC的干性。弗雷德·哈钦森癌症研究中心P35。Daniel E. Kennedy DNMT3A功能丧失突变会损害感染期间贝勒医学院p36期间免疫记忆和先天细胞效应功能的发展。Alana M. Franceski芯片相关的外在因素,塑造健康的造血干细胞O'Neal综合癌症中心p37。Emily Tsao通过STAU1损失的转录后调节有助于DEL(20q)无序的造血性造血中心玛格丽特玛格丽特癌症中心和多伦多大学P38的造血细胞分化缺陷。patrick Stelmach突变特异性表型DNMT3A突变干细胞在克隆造血中心德国癌症研究中心Alexander Marr BRD4抑制作用在TET2突变的克隆造血的鼠模型中消除了炎症和自我更新。Alexander Marr BRD4抑制作用在TET2突变的克隆造血的鼠模型中消除了炎症和自我更新。

产品表Neuro-2a细胞| 400394
N2A细胞系还提供了有关各种基因和蛋白质在神经元功能和发育中的作用的见解。例如,已在神经2A细胞中研究了以其参与DNA甲基化过程而闻名的DNMT3A基因,以了解其对神经元细胞和神经发育过程的影响。这些细胞中人甲状腺激素受体的表达使研究人员可以研究甲状腺激素反应及其对神经发育的影响以及神经母细胞瘤细胞的影响为更成熟的神经元表型。蛋白激酶信号通路是N2A细胞中强烈研究的另一个领域,鉴于它们在介导各种细胞过程(包括细胞生长,分化和对细胞外信号的反应)中的关键作用。蛋白激酶信号通路是N2A细胞中强烈研究的另一个领域,鉴于它们在介导各种细胞过程(包括细胞生长,分化和对细胞外信号的反应)中的关键作用。

产品表CélulasHCC78 | 302156
N2A细胞系还提供了有关各种基因和蛋白质在神经元功能和发育中的作用的见解。例如,已在神经2A细胞中研究了以其参与DNA甲基化过程而闻名的DNMT3A基因,以了解其对神经元细胞和神经发育过程的影响。这些细胞中人甲状腺激素受体的表达使研究人员可以研究甲状腺激素反应及其对神经发育的影响以及神经母细胞瘤细胞的影响为更成熟的神经元表型。蛋白激酶信号通路是N2A细胞中强烈研究的另一个领域,鉴于它们在介导各种细胞过程(包括细胞生长,分化和对细胞外信号的反应)中的关键作用。蛋白激酶信号通路是N2A细胞中强烈研究的另一个领域,鉴于它们在介导各种细胞过程(包括细胞生长,分化和对细胞外信号的反应)中的关键作用。