XiaoMi-AI文件搜索系统
World File Search SystemDSB

丁酰乳糖A是一种磷脂氟脂肪抑制剂,可增强
修复DNA损伤对于所有生物体来说都是至关重要的。DNA双链断裂(DSB)是最严重的DNA损伤类型之一,因为它们导致丧失了网络信息和未修复时死亡。在大肠杆菌中,它们被RECBCD复合物认识和处理,该复合物通过同源重组启动修复。尽管RECBCD下游的重复动力学已得到很好的特征,但尚不清楚该复合物与DNA保持附着多长时间,以及什么触发了其在体内的分离。要回答这些问题,我们在单分子水平上成像了RECB,并量化了其在暴露于环丙沙星的细菌细胞中的动态行为,这是一种诱导DSB的抗生素。我们的结果表明,RECB与DSB(10秒)形成长寿命的复合物,并且其与DNA的解离是复合物的固有证券,不取决于DNA损伤的量,也不取决于修复途径中的以下步骤。更重要的是,我们表明我们可以使用与DSB的RECB结合作为估计损害形成速率的标记。这项研究对RECBCD与DNA双链在体内的大肠杆菌的相互作用以及对环丙沙星诱导的DSB的细菌反应提供了详细的定量见解。

通过DNA聚合酶辅助的终端标记
在彗星测定中的摘要中,如果细胞被X X倍化为Genoto XIC剂,则在单细胞凝胶电泳后形成尾巴。these尾巴包括DNA单链断裂(SSB)和双链断裂(DSB)的混合物。ho w e v er,这些两种类型的链断裂无法使用具有Con V en ventionDNA染色的彗星测定方案来区分。由于DSB对单元格是有问题的,因此如果可以在同一彗星中差异化SSB和DSB,则将很有用。为了能够区分SSB和DSB,我们为聚合酶辅助的DNA损伤分析(PADDA)设计了一种协议,可与Flash Comet协议或固定单元格结合使用。通过使用DNA聚合酶I将SSB和末端脱氧核苷酸转移酶标记为具有荧光团标记的核苷酸的DSB。在此,TK6细胞或HACAT细胞暴露于过氧化氢(H 2 O 2),电离辐射(X射线)或DNA切割酶,然后遵循彗星方案,以实施彗星方案。p adda提供了更广泛的检测范围,未发现的DNA链断裂的未发现的未发现。
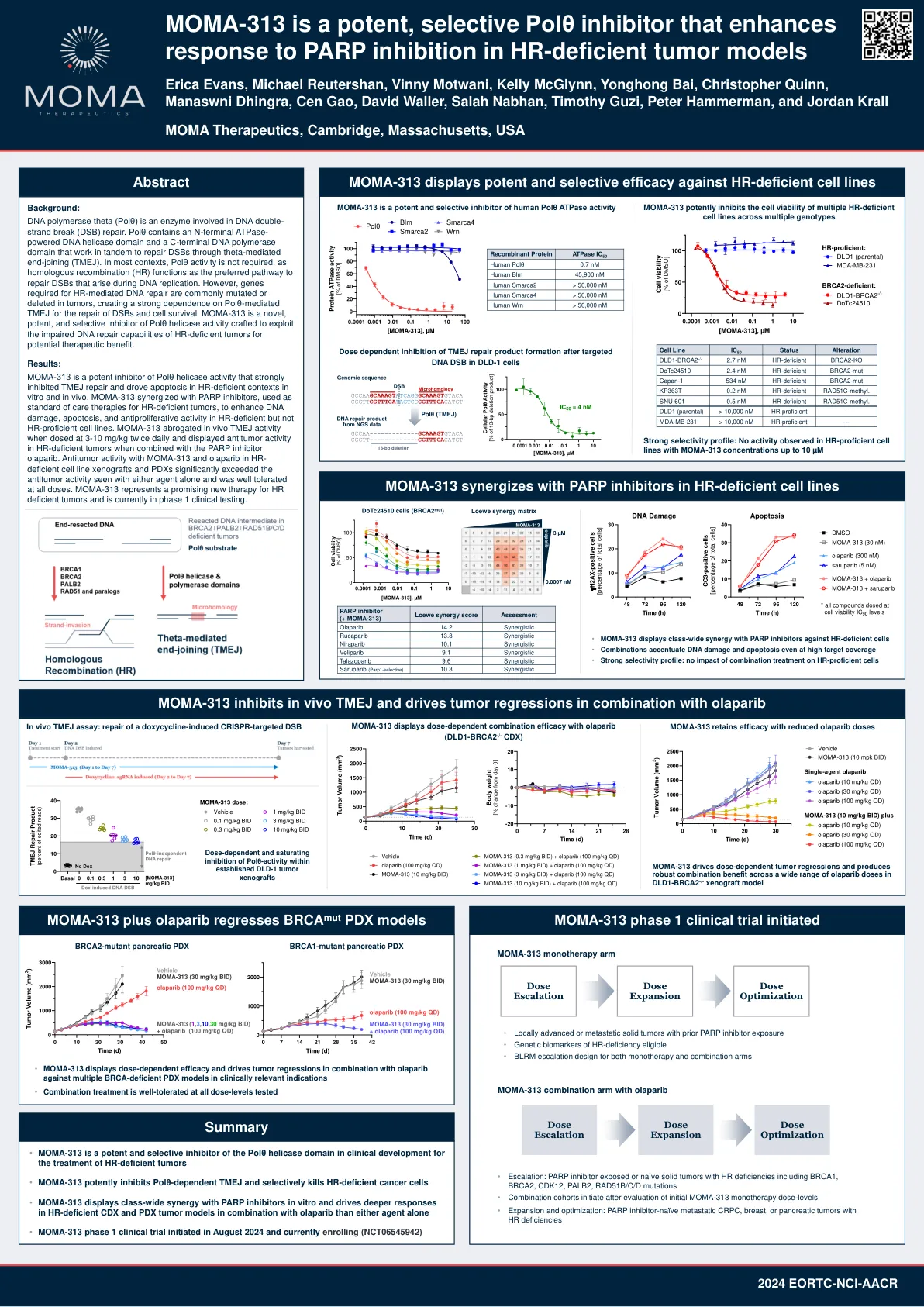
MOMA-313 是一种强效、选择性 Polθ 抑制剂,可增强 HR 缺陷肿瘤模型对 PARP 抑制的反应
DNA 聚合酶 theta (Polθ) 是一种参与 DNA 双链断裂 (DSB) 修复的酶。Polθ 包含一个 N 端 ATPase 驱动的 DNA 解旋酶结构域和一个 C 端 DNA 聚合酶结构域,它们协同作用,通过 theta 介导的末端连接 (TMEJ) 修复 DSB。在大多数情况下,Polθ 活性不是必需的,因为同源重组 (HR) 是修复 DNA 复制过程中出现的 DSB 的首选途径。然而,HR 介导的 DNA 修复所需的基因通常在肿瘤中发生突变或缺失,导致 DSB 修复和细胞存活严重依赖 Polθ 介导的 TMEJ。MOMA-313 是一种新型、有效且选择性的 Polθ 解旋酶活性抑制剂,旨在利用 HR 缺陷型肿瘤受损的 DNA 修复能力来获得潜在的治疗益处。

集成传感器收集的情报
本报告是国防科学委员会 (DSB) 的产物,并根据所有联邦咨询委员会法案 (FACA) 法规发布。DSB 是一个联邦咨询委员会,旨在为国防部长提供独立建议。本报告中的声明、意见、结论和建议不一定代表国防部的官方立场。情报科学委员会 (ISB) 就情报界特别重要的新兴科学和技术问题向国家情报总监办公室 (ODNI) 和高级情报界领导人提供建议。本报告中的声明、意见、结论和建议不一定代表情报界任何机构的官方立场。DSB/ISB 传感器收集情报整合联合工作组于 2008 年 1 月完成了信息收集工作。本报告为非机密报告,可向公众发布。

使用 CRISPR 的有针对性的、可调节的 DNA 损伤工具......
摘要:哺乳动物细胞不断受到各种 DNA 损伤事件的影响,从而导致 DNA 修复途径的激活。了解 DNA 损伤反应的分子机制有助于开发针对这些途径元素的治疗方法。双链断裂 (DSB) 对细胞活力和基因组稳定性特别有害。通常,使用 DNA 损伤剂(例如电离辐射或基因毒性药物)研究 DSB 修复。这些会在难以控制损伤剂量的非预测基因组位点处引起随机损伤。此类干预不适合研究特定 DSB 位点如何根据局部染色质状态调用不同的 DNA 损伤识别和修复途径。RNA 引导的 Cas9(CRISPR 相关蛋白 9)核酸内切酶是介导靶向基因组改变的强大工具。基于 Cas9 的基因组干预是通过在感兴趣的基因组区域形成 DSB 来实现的。在这里,我们利用基于计算机预测的定制设计的混杂向导 RNA,在人类基因组的特定数量和位置诱导 DSB。这是通过重组 Cas9-向导复合物的电穿孔实现的,它提供了一种通用、低成本且快速的方法,用于在细胞培养模型中诱导受控 DNA 损伤。

通过靶向组蛋白 H2A-K15 泛素化增强 CRISPR-Cas9 诱导的精准基因编辑
修改,例如疾病突变建模或体细胞基因治疗中突变的校正。为了加强精确的基因编辑,需要工具或干预措施使 DSB 修复途径选择偏向 HDR,并通过将 DNA 修复模板靶向递送到 DSB 来促进 HDR 处理。特别是修复模板的可用性可能是 HDR 的限速因素。以前用于靶向递送修复模板的方法使用 Cas9 融合蛋白,其结构域与功能基团结合,该功能基团被掺入合成寡核苷酸或 PCR 片段中作为供体模板,并作为组合的 Cas9-sgRNA-供体复合物递送到细胞中 [3-5]。然而,目前尚不清楚修复模板分子与 Cas9 核酸酶的连接是否是共递送的最有效方式,因为模板在 DSB 修复的后续步骤中是必需的。以前促进 DSB 修复途径选择的方法有利于

ßA1-晶体蛋白 HDR 质粒 (m): sc-419822-HDR
含有由 CRISPR/Cas9 系统产生的双链断裂 (DSB) 的 DNA 可以通过非同源末端连接 (NHEJ) 或同源定向修复 (HDR) 途径进行修复 (1,2,3)。NHEJ 修复途径在切割位点引入非特异性插入或缺失,而 HDR 途径允许在 DSB 位点进行精确的基因编辑 (1,2,3)。靶向特异性 HDR 质粒为 DSB 提供 DNA 修复模板,当与 CRISPR/Cas9 KO 质粒共转染时,能够在发生 Cas9 诱导的 DNA 切割的位置插入特定的选择标记 (1,2)。HDR 质粒可以整合红色荧光蛋白 (RFP) 基因以直观地确认转染,并整合抗生素抗性基因 (嘌呤霉素) 以选择含有成功 CRISPR/Cas9 双链断裂的细胞。嘌呤霉素抗性和 RFP 编码基因两侧是两个 LoxP 位点,这些位点可被 Cre 载体识别,之后可利用该位点从基因组 DNA 中去除这些选择标记 (4,5)。

染色体修复辅助途径在酵母中改组
图1 - 酵母中染色体修复(CR)和外源修复(ER)途径的比较。A,ER和CR路径的概述。Cas9介导的DSB可以通过外源或染色体供体修复。er会导致外源供体的整合,而CR会重复现有的染色体供体。b,ER和CR途径的修复效率。使用一个ER供体或越来越多的CR供体引入和修复了CAS9 DSB。修复模板(LPA-REN-LPZ)旨在将LPA-T9-LPZ的CAS9目标位点突变为限制性核酸内切酶识别序列(REN),以促进筛选。对照显示在没有修复模板(无修复)和没有GRNA(无DSB)的情况下描述生存能力。生存能力(已修复的DSB的比例)。错误条代表S.D.三个生物学重复。c,不同大小的CR同源性区域的CR效率。cr模板具有从60 bp到280 bp的长度不等的同源区域,并将其整合到同一染色体基因座中,并用于修复Cas9 DSB。cr生存能力 1B。 错误条代表S.D. 三个生物学重复。1B。 错误条代表S.D. 三个生物学重复。1B。错误条代表S.D.三个生物学重复。

DNA双链断裂修复的新方面
摘要:放射治疗是当今癌症管理的重要组成部分,利用不同方式的电离辐射(IR)来减轻癌症的进展。ir功能。其中最有害的是DNA双链断裂(DSB)。在进化过程中,较高的真核生物的细胞已经发展出四个主要的DSB修复途径:经典的非同源末端连接(C-NHEJ),同源重组(HR),替代性最终连接(ALT-EJ)(ALT-EJ)和单链退火(SSA)。这些机械上不同的修复途径具有不同的细胞周期和同源性依赖性,但令人惊讶的是,它们具有截然不同的效果和动力学的作用,因此对细胞存活和基因组维持无效。因此,在这些DSB修复途径的参与中预期进行严格的调节和协调是合理的,以实现最大可能的基因组稳定性。在这里,我们提供了有关这些修复途径支撑的分子机制的累积知识的最新综述,重点是C-NHEJ和HR。我们讨论了最近出现的因素和过程。我们概述了整个细胞周期中DSB修复途径选择的机制,并突出了DNA终端切除在此过程中的关键作用。然而,最重要的是,我们指出在低DSB载荷下对HR的强烈偏好,因此对于在细胞周期的G 2期中受辐照的细胞而言,IR剂量较低。我们进一步探讨了从高层到低限制误差的修复途径的过渡的分子基础,并分析了这种过渡对细胞生存能力和基因组稳定性的协调和后果。最后,我们详细阐述了这些进步如何有助于制定放射治疗中的癌症治疗方案。

CRISPR/CAS9的基因组编辑工具箱,用于拟南芥
CRISPR/CAS9系统已成为一种强大的基因组工程工具,用于研究基因功能并改善植物特征。基因组编辑是通过Cas9核酸内切酶在特定的基因组序列上实现的,以产生由短导RNA(SGRNA)指导的双标准断裂(DSB)。DSB通过容易出错的非同源末端连接(NHEJ)或无错误的同源指导修复(HDR)路径来修复,分别导致基因突变或序列替换。这些细胞DSB修复途径可以被利用以敲除或替换基因。另外,胞质或腺嘌呤碱基编辑器(CBES或ABE)融合到催化死亡的Cas9(DCAS9)或Nickase Cas9(NCAS9)(NCAS9)时,也用于执行精确的基础编辑而无需生成DSB。在本章中,我们描述了通过使用基于CRISPR/CAS9的系统在拟南芥基因组中执行单个/多基因突变和精确基础编辑的详细程序。特别是,描述了转基因线的目标基因选择,SGRNA设计,矢量结构,转化和分析的步骤。该方案有可能适应在其他植物物种(例如水稻)中进行基因组编辑。