XiaoMi-AI文件搜索系统
World File Search SystemFEN1

C3 / Padres Pedal the Cause 2020“FEN1 核酸酶 - ...
尤文氏肉瘤 (ES) 是由致癌的 EWS-FLI1 融合蛋白引起的,该融合蛋白是由 t (11; 22) 染色体易位引起的。EWS-FLI1 将 BRCA1 隔离在转录复合物中,以防止其进行同源重组 (HR)。与这种 HR 缺陷一致,癌细胞系药物敏感性项目发现 ES 细胞对 PARP 抑制剂敏感。HR 是抑制 DNA 复制错误以维持基因组完整性的众多途径之一。酵母遗传学已经阐明了包括 HR 在内的途径之间的合成致死关系,这些关系可以解决停滞的复制叉,并且可以作为癌症治疗的靶点。在酵母中,在滞后链 DNA 合成中起作用的 RAD27(人类 FEN1)核酸内切酶的突变与 HR 基因的突变一起是合成致死的。我们已经证明,这种合成致死关系是保守的,因为人类 BRCA 突变癌细胞对 FEN1 抑制高度敏感。有趣的是,我们发现 PARP 抑制剂抗性的 ES 细胞系 (SK-ES-1) 对我们专有的 FEN1 抑制剂的敏感性与 BRCA 突变癌细胞一样。通过挖掘大规模 CRIPSR 筛选数据库,我们发现 FEN1 对所有五种测试的 ES 细胞系都具有独特的必要性。为了评估 FEN1 作为 ES 治疗的潜在靶点,该合作试点项目将进行三项研究:1) 将 FEN1 抑制剂结果扩展到一组 ES 细胞系,并与 PARP 抑制剂和临床相关化疗药物进行比较研究;2) 使用 siRNA 敲低 FEN1 来验证 FEN1 抑制剂研究;3) 确定 EWS-FLI1 融合蛋白如何促进 ES 细胞对 FEN1 的依赖。

FEN1 内切酶作为同源重组缺陷人类癌症的治疗靶点
癌症治疗的合成致死策略利用癌症特异性基因缺陷来识别对肿瘤细胞存活至关重要的靶点。本文我们表明,RAD27/FEN1 编码瓣状内切酶 1 (FEN1),这是一种在 DNA 复制和修复中发挥作用的结构特异性核酸酶,与酿酒酵母基因组不稳定性基因具有最多的合成致死相互作用,是基于抑制剂的方法杀死同源重组 (HR) 缺陷癌症的可用药物靶点。研究表明,小分子 FEN1 抑制剂和 FEN1 小干扰 RNA (siRNA) 可选择性杀死 BRCA1 和 BRCA2 缺陷的人类细胞系,从而证实了 HR 缺陷癌症容易受到 FEN1 缺失的影响。此外,在小鼠中重现了对 FEN1 抑制的不同敏感性,小分子 FEN1 抑制剂降低了药物敏感但无耐药性癌细胞系中形成的肿瘤的生长。FEN1 抑制在敏感和耐药细胞系中均诱导了 DNA 损伤反应;然而,即使去除抑制剂,敏感细胞系也无法恢复和复制 DNA。尽管 FEN1 抑制在敏感细胞中将 caspase 激活到更高水平,但这种凋亡反应发生在 p53 缺陷细胞中,而泛 caspase 抑制剂无法阻断细胞杀伤。这些结果表明,FEN1 抑制剂具有治疗靶向 HR 缺陷癌症的潜力,例如由 BRCA1 和 BRCA2 突变和其他遗传缺陷引起的癌症。
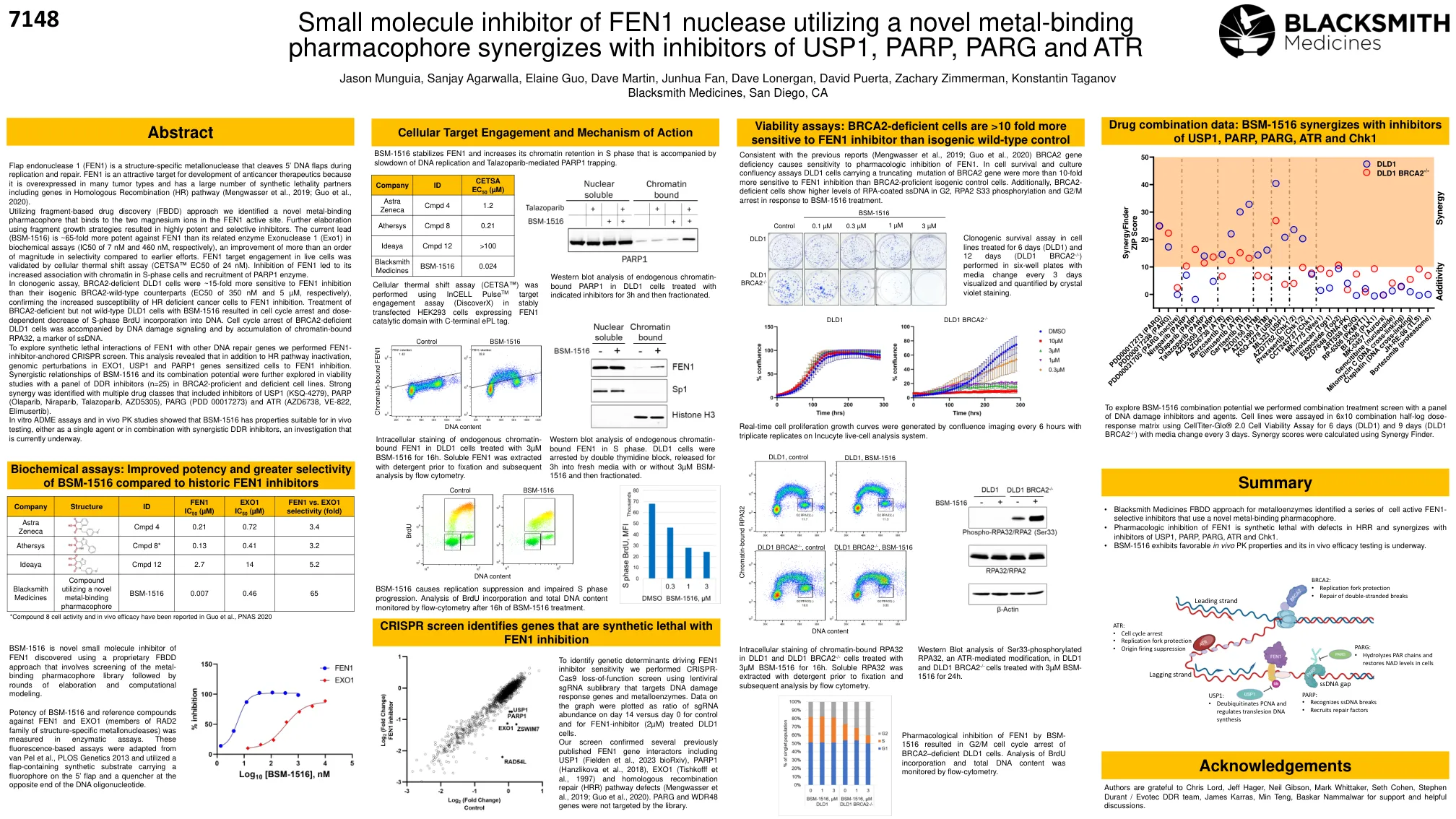
使用新型金属结合药效团与USP1,PARP,PARG和ATR抑制剂协同的Fen1核酸酶的小分子抑制剂
皮瓣核酸内切酶1(Fen1)是一种结构特异性的金属核酸酶,在复制和修复过程中切割5'DNA瓣。fen1是开发抗癌疗法的有吸引力的靶标,因为它在许多肿瘤类型中过表达,并且具有大量的合成致死性伴侣,包括同源重组基因(HR)途径(Mengwasser等,2019; Guo等,2020)。利用基于碎片的药物发现(FBDD)方法,我们确定了一种新型的金属结合药效团,该药效团与Fen1活性位点中的两个镁离子结合。使用碎片增长策略进一步阐述导致高度有效和选择性抑制剂。在生物化学测定中(分别为7 nm和460 nm的IC50),对FEN1的当前铅(BSM-1516)对FEN1的有效性比其相关酶外核酸酶1(EXO1)高65倍,与早期努力相比,改善了量的更大范围。fen1靶标在活细胞中的靶标参与通过细胞热偏移分析验证(CETSA

flap核酸内切酶1通过ADP-核糖基化Yilun Sun 1*,Lisa M. Jenkins 2,Lara H. El Touny 3,Ukhyun Jo 1,Xi Yan
抽象的DNA-蛋白交联(DPC)是最普遍和有害的DNA病变之一,是由于暴露于代谢应激,药物或交联药物(如甲醛(FA))而引起的。fa是甲醇代谢,组蛋白脱甲基化,脂质过氧化和环境污染物的细胞副产品。无法修复FA诱导的DPC几乎所有基于染色质的过程,包括复制和转录,导致免疫缺陷,神经变性和癌症。然而,它在很大程度上仍然未知细胞如何维修DPC。由于缺乏鉴定DPC的技术,我们不理解FA的蛋白质类型会阻碍DPC修复的研究。在这里,我们通过将氯化葡萄球菌差异超速离心与HPLC-MAS-MAS光谱法(MS)耦合,从而设计了一种新型的生物测定法,以介绍FA诱导的DPC。使用该方法,我们揭示了FA诱导的人类细胞中FA诱导的DPC的蛋白质组,发现形成DPC的最丰富的蛋白质是PARP1,拓扑异构酶I和II和II和II,甲基转移酶,DNA和RNA聚合酶,组蛋白,组蛋白,以及核糖体蛋白。为了鉴定修复DPC的酶,我们进行了RNA干扰筛选,发现皮瓣核酸内切酶1(FEN1)的下调使细胞对FA过敏。由于Fen1具有5'-FLAP内切酶活性,因此我们假设FA诱导了DPC偶联的5'-FLAP DNA片段,可以通过Fen1处理。的确,我们证明了FA会损坏通过碱基切除途径(BER)转化为5'-FLAP的DNA碱基。我们还观察到受损的DNA碱基与DPC和FEN1共定位。从机械上讲,我们显示了FEN1在体内修复FA诱导的DPC和裂解5'-FLAP DNA底物,这些DNA具有模拟于体外的DPC。我们还发现,FEN1修复酶拓扑异构酶II(TOP2)-DPC,由其抑制剂依托泊苷和阿霉素诱导的诱导的酶促蛋白酶和阿霉素独立于BER途径,而FEN1和FEN1和DPC靶向的蛋白酶sprtn是对两种FA诱导的非Zym Zym Zym Zymations sprapterations spr的可行途径top2-dpcs。值得注意的是,我们发现FA诱导的非酶DPC和酶ToP2-DPC迅速通过聚辅助核糖基化(ParyLation)迅速修饰,这是一种由PARP1催化的翻译后修饰,由PARP1催化的,这是一种由Paryling DNA损伤损害蛋白和DNA Reparion Reparte resation and DNA损伤蛋白的关键DNA损伤效应器和DNA Reparte resation and dna Reparte stotes和DNA Reparte stotes。,我们用HPLC-MS的抗PAR抗体进行了免疫沉淀(IP)测定,并将Fen1鉴定为parylation底物。接下来,我们表明DPC底物的填充信号发出了Fen1,而Fen1的抚养也将Fen1驱动到DPC位点。最后,使用末端ADP-ribose-MS方法的酶促标记,我们将FEN1的E285残基确定为主要的荷置位点,这似乎是FEN1迁移到DPCS所需的。综上所述,我们的工作不仅揭示了FA诱导的DPC的身份,而且还发现了前所未有的PARP1-FEN1核酸酶途径,是一种通用和势在必行的机制,可以修复其他DPC并防止DPC诱导的基因组不稳定。

PARG 缺陷的肿瘤细胞对 EXO1/FEN1 介导的 DNA 修复的依赖性增强
目前正在研究以聚(ADP-核糖)糖基水解酶 (PARG) 为靶点治疗各种癌症,但我们对导致癌细胞易受这种定制疗法影响的特定遗传弱点了解甚少。此外,识别此类弱点对于靶向 BRCA2;p53 缺陷型肿瘤很有意义,这些肿瘤通过 PARG 表达丧失而获得对聚(ADP-核糖)聚合酶抑制剂 (PARPi) 的耐药性。在这里,通过进行全基因组 CRISPR/Cas9 缺失筛选,我们识别出参与 DNA 修复的各种基因,这些基因对于 PARG;BRCA2;p53 缺陷型细胞的存活至关重要。特别是,我们的研究结果揭示了 EXO1 和 FEN1 是 PARG 缺失的主要合成致死相互作用因子。我们提供了证据表明,在 PARG;BRCA2;p53 缺陷细胞中,复制叉进展、DNA 单链断裂修复和冈崎片段处理受损,这些改变加剧了 EXO1/FEN1 抑制的效果,并在这种情况下变得致命。由于这种敏感性取决于 BRCA2 缺陷,我们建议在失去 PARG 活性的 PARPi 抗性肿瘤中靶向 EXO1/FEN1。此外,EXO1/FEN1 靶向可能是增强 PARG 抑制剂在同源重组缺陷肿瘤中效果的有效策略。

tlr-2和
摘要遗传物质的稳定性和完整性对于维持和延续生活至关重要。人类基因组由三十亿对碱基组成,编码30,000-40,000个基因,并不断受到内源性反应性代谢产物,治疗药物和众多影响其完整性的环境诱变药物的攻击。因此,很明显,基因组的稳定性必须在连续监测之下。这是通过DNA修复机制实现的,DNA修复机制已开发出来去除或耐受DNA损伤和误差。在生物体中存在的DNA修复机制中,它们可以分为:i)基础切除修复(BER),ii)核苷酸切除(NER),iii)基本MALPASE(MMR)和IV)DNA修复,通过非同型末端(NHEJ)。对于这些机制正常工作,很明显,负责修复功能的蛋白质之间相互作用的重要性,以及对负责提到的机制的蛋白质正确位置的核进口调节。在负责调节核进口的机制中,由进口异二聚体α/β组成的经典途径是位移的主要机制之一。某些修复蛋白似乎仅与进口α(IMP)的某些同样蛋白相互作用,表明对修复过程的额外调节,但对这些蛋白质的核位置序列(NLS)的识别知之甚少。通过这些结果,阐明了包含NLSS KU80和FEN1的结构。这项工作特别涉及使用晶体学技术与蛋白质相关DNA修复的NLSS肽的IMP复合物的结构复合物的研究。进行了在其N末端部分截断的Musculus印象的表达和纯化,以及与DNA相关蛋白的NLS肽的IMP偶然化,对应于KU80,PMS2和MLH1蛋白质和MLH1蛋白质和BIPARTARTARTARTATTITE序列的单型序列。X射线衍射数据,并以2.1-2.38Å的分辨率进行处理。肽NLS KU80与NHEJ修复有关,与主连接位点上的IMPα相似,类似于SV40 T抗原的NLS(S 1)。已经与ber修复有关的NLS FEN1肽与Sitia S 1和次级位点(S 2)有关,证明是两部分序列。此外,仅具有10种废物的Fen1肽接头区域使与IMPα的联系更好,并且与具有11-12废物的肽的连接相比,与IMPα的连接更扩展,可能更有利的构象。在连接位点上的特定位置被确认为必不可少的,以及在这些区域中保守的残留物,表明这些位点中分子间相互作用的重要性。此信息表明这些蛋白质可以通过IMP-α独立运输到核心,而无需与有关修复的其他蛋白质形成复合物。关键字:进口α,核进口,NLS,射线晶体学-X,KU80,FEN1,PMS2,MLH1。

教学:分子生物学
基本所有者程序。分子生物学研究领域。<生物学的女主角教条。分子生物学中最常用的测量单元。c ristalloghich to x -rays和分子建模。x体晶体学。van der waals基于射线的模型。溶剂表面和浅表静电电位。氢桥线的结构几何形状。c核酸的结构射流。核苷和核苷酸。 磷酸化的脑结合和主要结构。 DNA二级结构。 DNA B和DNA A. RNA的二级和三级结构的结构参数。 基因组对DNA的 r恢复。 Meselson和Stahl实验。 冈崎的碎片。 大肠杆菌中的复制:首先,DNA聚合酶III,DNA聚合酶I,DNA Ligasi。 真核染色体DNA的复制:DNA Polimerasi alfa,DNA Polimerasi Delta,ribonucleasi H,Endonucleasi fen1。 人性线粒体DNA的复制。 端粒的作用。 的移动RNA的理解和成熟。 操纵子。 促进mRNA的结构。 RNA均值聚合酶和相对启动子。 cappuccio组。核苷和核苷酸。磷酸化的脑结合和主要结构。DNA二级结构。DNA B和DNA A. RNA的二级和三级结构的结构参数。 基因组对DNA的 r恢复。 Meselson和Stahl实验。 冈崎的碎片。 大肠杆菌中的复制:首先,DNA聚合酶III,DNA聚合酶I,DNA Ligasi。 真核染色体DNA的复制:DNA Polimerasi alfa,DNA Polimerasi Delta,ribonucleasi H,Endonucleasi fen1。 人性线粒体DNA的复制。 端粒的作用。 的移动RNA的理解和成熟。 操纵子。 促进mRNA的结构。 RNA均值聚合酶和相对启动子。 cappuccio组。DNA B和DNA A. RNA的二级和三级结构的结构参数。基因组对DNA的 r恢复。 Meselson和Stahl实验。 冈崎的碎片。 大肠杆菌中的复制:首先,DNA聚合酶III,DNA聚合酶I,DNA Ligasi。 真核染色体DNA的复制:DNA Polimerasi alfa,DNA Polimerasi Delta,ribonucleasi H,Endonucleasi fen1。 人性线粒体DNA的复制。 端粒的作用。 的移动RNA的理解和成熟。 操纵子。 促进mRNA的结构。 RNA均值聚合酶和相对启动子。 cappuccio组。r恢复。Meselson和Stahl实验。冈崎的碎片。大肠杆菌中的复制:首先,DNA聚合酶III,DNA聚合酶I,DNA Ligasi。真核染色体DNA的复制:DNA Polimerasi alfa,DNA Polimerasi Delta,ribonucleasi H,Endonucleasi fen1。人性线粒体DNA的复制。端粒的作用。的移动RNA的理解和成熟。操纵子。促进mRNA的结构。RNA均值聚合酶和相对启动子。cappuccio组。转录和多掺杂终止。内含物和剪接。RNA编辑。 Matui真核mRNA结构。 遗传密码。 RNA中基因组的 r。 pury-极性RNA复制机制(黄病毒,picornavirus,逆转录病毒),阴性极性RNA病毒,双丝细丝RNA病毒。 肝病病毒的特殊性。 的理解蛋白质。 运输RNA的结构和功能。 tRNA氨基acancezion。 <核糖体的分裂结构和功能特征。 将转化为过程和真核生物的开始。 <分配扩展翻译的阶段。 翻译的终止。 发射。 阅读阶段的滑动。 基因组序列的Nterpotation。 原核生物和真核编码基因的典型结构。 鉴定开放阅读方案(ORF),基因表达控制的内含子和元素。 基因表达的 r抑制。 调整了Procarials中转录开始的开始:组成型控制和调节控制。 真核生物中转录开始的开始。 家政和特定于织物的基因。 <结合DNA的蛋白质的分裂结构基序:螺旋螺旋螺旋,锌指,亮氨铰链。RNA编辑。Matui真核mRNA结构。遗传密码。RNA中基因组的 r。 pury-极性RNA复制机制(黄病毒,picornavirus,逆转录病毒),阴性极性RNA病毒,双丝细丝RNA病毒。 肝病病毒的特殊性。 的理解蛋白质。 运输RNA的结构和功能。 tRNA氨基acancezion。 <核糖体的分裂结构和功能特征。 将转化为过程和真核生物的开始。 <分配扩展翻译的阶段。 翻译的终止。 发射。 阅读阶段的滑动。 基因组序列的Nterpotation。 原核生物和真核编码基因的典型结构。 鉴定开放阅读方案(ORF),基因表达控制的内含子和元素。 基因表达的 r抑制。 调整了Procarials中转录开始的开始:组成型控制和调节控制。 真核生物中转录开始的开始。 家政和特定于织物的基因。 <结合DNA的蛋白质的分裂结构基序:螺旋螺旋螺旋,锌指,亮氨铰链。r。pury-极性RNA复制机制(黄病毒,picornavirus,逆转录病毒),阴性极性RNA病毒,双丝细丝RNA病毒。肝病病毒的特殊性。的理解蛋白质。运输RNA的结构和功能。tRNA氨基acancezion。<核糖体的分裂结构和功能特征。将转化为过程和真核生物的开始。<分配扩展翻译的阶段。翻译的终止。发射。阅读阶段的滑动。基因组序列的Nterpotation。原核生物和真核编码基因的典型结构。鉴定开放阅读方案(ORF),基因表达控制的内含子和元素。r抑制。调整了Procarials中转录开始的开始:组成型控制和调节控制。真核生物中转录开始的开始。家政和特定于织物的基因。<结合DNA的蛋白质的分裂结构基序:螺旋螺旋螺旋,锌指,亮氨铰链。染色质结构对基因表达的影响:组蛋白的乙酰化和扩展; DNA甲基化。由microRNA介导的天才沉默。<用于分析核酸的Diva Basic etohs。紫外光谱和量化

源自CRISPR-CAS I-版本的系统...
主要版本(PE)保留了CRISPR的特定靶向靶向,但以RNA模型的形式采用了额外的货物,其中包含修改作为导向RNA(称为PEGARN)的连续估计。要求修饰蛋白质的情况,以使Cas9(H840A)仅裂解,而且还需要关联(PE1),或在其C端(PE2)合并与逆转录酶M-MLV(RT)(RT)(H840A)结束。使用Cas9(H840a)的使用(通常称为Nickase Cas9)避免形成双链DNA断裂(DSB),并简单地切割了PAM位点上游的DNA的非全面链。该表现出具有OH 3'基团的DNA瓣,该小组结合了RNA矩阵的引物(PBS)的联络位点,用作RT的底漆,该引物通过复制Pegarn的版本序列来扩展襟翼3'。尽管在热力学上,与5'未出版的皮瓣相比,杂交未发表的互补链的可能性较小,但内源性内核酸内核酸酶Fen1的固有偏好是消除5'碎片,导致3'编辑皮瓣的杂交导致了非常有效的基本版本。

片段 - 印第安纳大学印第安纳波利斯分校 ScholarWorks
癌症将直接影响超过三分之一人口的生活。DNA损伤反应 (DDR) 是一个复杂的系统,涉及损伤识别、细胞周期调控、DNA 修复以及最终的细胞命运决定,在癌症病因和治疗中发挥着核心作用。涉及 DDR 靶向的两种主要治疗方法包括:采用抗癌基因毒性剂的组合疗法;以及合成致死,利用散发性 DDR 缺陷作为癌症特异性治疗的机制。尽管许多 DDR 蛋白已被证明“无法用药”,但基于片段和结构的药物发现 (FBDD、SBDD) 已推进了治疗剂的鉴定和开发。FBDD 已促成 4 种药物(另有约 50 种药物处于临床前和临床开发阶段),而据估计,SBDD 已促成 200 多种 FDA 批准药物的开发。基于蛋白质 X 射线晶体学的片段库筛选,特别是针对难以捉摸或“无法用药”的靶标,可以同时生成命中结果以及蛋白质-配体相互作用和结合位点(正构或变构)的详细信息,从而为化学可处理性、下游生物学和知识产权提供信息。使用一种新型的高通量基于晶体学的片段库筛选平台,我们筛选了五种不同的蛋白质,命中率约为 2 e 8%,晶体结构约为 1.8 至 3.2 Å。我们考虑了当前的 FBDD/SBDD 方法和一些设计针对 DDR 核酸酶减数分裂重组 11(MRE11,又名 MRE11A)、无嘌呤/无嘧啶核酸内切酶 1(APE1,又名 APEX1)和 flap 核酸内切酶 1(FEN1)的抑制剂的示例性结果。© 2020 作者。由 Elsevier Ltd. 出版。这是一篇根据 CC BY-NC-ND 许可协议 ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ) 开放获取的文章。

分段
癌症将直接影响超过三分之一人口的生活。DNA损伤反应 (DDR) 是一个复杂的系统,涉及损伤识别、细胞周期调控、DNA 修复以及最终的细胞命运决定,在癌症病因和治疗中发挥着核心作用。涉及 DDR 靶向的两种主要治疗方法包括:采用抗癌基因毒性剂的组合疗法;以及合成致死,利用散发性 DDR 缺陷作为癌症特异性治疗的机制。尽管许多 DDR 蛋白已被证明“无法用药”,但基于片段和结构的药物发现 (FBDD、SBDD) 已推进了治疗剂的鉴定和开发。FBDD 已促成 4 种药物(另有约 50 种药物处于临床前和临床开发阶段),而据估计,SBDD 已促成 200 多种 FDA 批准药物的开发。基于蛋白质 X 射线晶体学的片段库筛选,特别是针对难以捉摸或“无法用药”的靶标,可以同时生成命中结果以及蛋白质-配体相互作用和结合位点(正构或变构)的详细信息,从而为化学可处理性、下游生物学和知识产权提供信息。使用一种新型的高通量基于晶体学的片段库筛选平台,我们筛选了五种不同的蛋白质,命中率约为 2 e 8%,晶体结构约为 1.8 至 3.2 Å。我们考虑了当前的 FBDD/SBDD 方法和一些设计针对 DDR 核酸酶减数分裂重组 11(MRE11,又名 MRE11A)、无嘌呤/无嘧啶核酸内切酶 1(APE1,又名 APEX1)和 flap 核酸内切酶 1(FEN1)的抑制剂的示例性结果。© 2020 作者。由 Elsevier Ltd. 出版。这是一篇根据 CC BY-NC-ND 许可协议 ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ) 开放获取的文章。