XiaoMi-AI文件搜索系统
World File Search SystemFoxp3
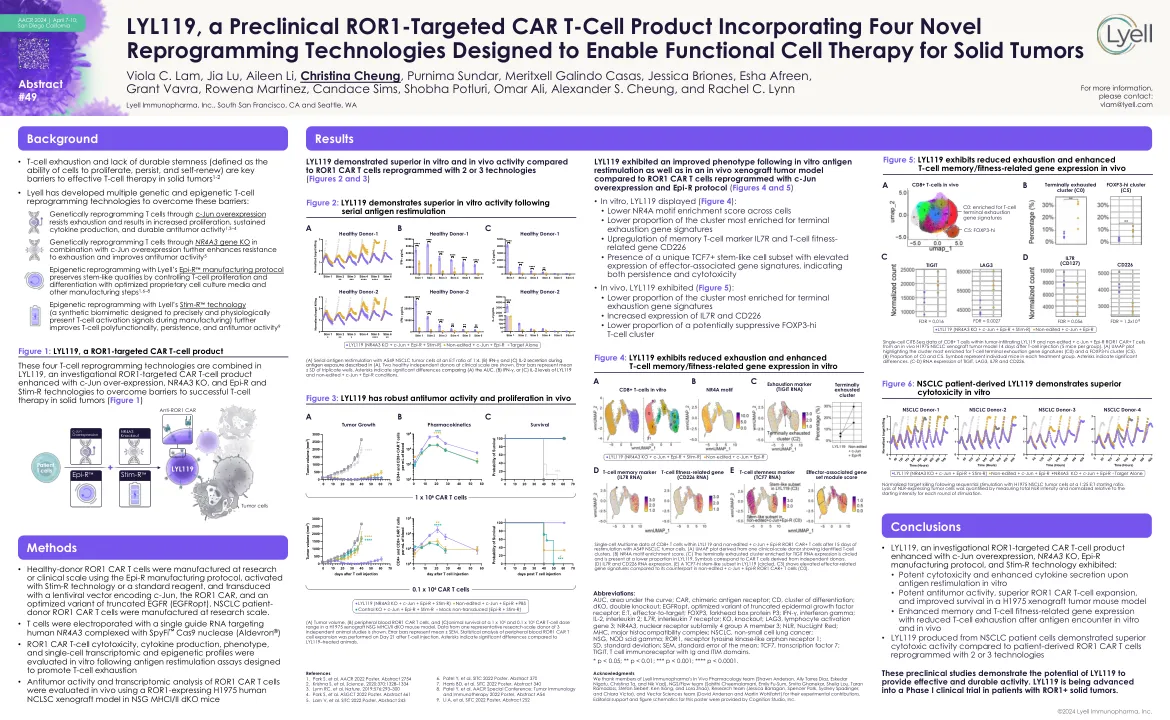
lyl119,一种临床前ROR1靶向的汽车T细胞产品,融合了四种新型重编程技术,旨在实现功能性细胞疗法
缩写:AUC,曲线下方的区域;汽车,嵌合抗原受体; CD,分化簇; DKO,双淘汰赛; EGFROPT,截短表皮生长因子受体的优化变体; e:t,效应器到目标; Foxp3,叉子盒蛋白P3; IFN-γ,干扰素伽玛; IL-2,白介素2; IL7R,白介素7受体; ko,淘汰; lag3,淋巴细胞激活基因3; NR4A3,核受体亚科4组成员3; NLR,Nutlight红色; MHC,主要的组织相容性复合物; NSCLC,非小细胞肺癌; NSG,点头SCID伽玛; ROR1,受体酪氨酸激酶样孤儿受体1; SD,标准偏差; SEM,平均值的标准误差; TCF7,转录因子7;带有IG和ITIM结构域的Tigit,T细胞免疫受体。

罕见免疫疾病为基因组编辑铺平道路......
成簇的规律间隔的短回文重复序列 (CRISPR) 基因组编辑平台预示着基因治疗新时代的到来。针对危及生命的血液和免疫系统单基因疾病的创新疗法正在从半随机基因添加转变为对缺陷基因的精确修改。随着这些疗法进入首次人体临床试验,它们的长期安全性和有效性将为未来一代基于基因组编辑的医学提供参考。在这里,我们讨论了先天性免疫缺陷作为建立和推进精准医疗的疾病原型的重要性。我们将回顾基于成簇的规律间隔的短回文重复序列的基因组编辑平台修改原代细胞 DNA 序列的可行性,并描述两种新兴的基因组编辑方法来治疗 RAG2 缺陷(一种原发性免疫缺陷)和 FOXP3 缺陷(一种原发性免疫调节障碍)。

TCF-1的补充信息促进了对异常β-catenin激活的反应,促进了基因组不稳定性和T细胞转化。 Stephen Arno
胸腺细胞在流式细胞仪缓冲液(PBS中的2%FBS)中表面染色30分钟。样品,并在LSRII流式细胞仪(Becton Dickinson)上获取数据。使用FlowJo软件(Becton Dickinson)分析数据。表面抗体是CD4(克隆GK1.5,BD Biosciences),CD8(克隆53-6.7,Ebiosciences),TCRβ(克隆H57-597,Ebiiosciences)和CCR7(克隆H57-597,CCR7(Clone 4B12,Ebiosciences,Ebiosciences)。细胞使用活/死水荧光反应性染料(分子探针与生命技术,L34963)染色。对于γH2AX实验,根据制造商的建议,将细胞固定并使用FOXP3/转录因子固定试剂盒(EBISoscience 00-5521)进行通透。细胞对γH2AX(抗H2AX(PS139),BD Biosciences,BDB562377)的细胞内染色30分钟,在冰川化缓冲液中冰上进行30分钟,洗涤2倍,并获得上述收购。

疟疾中的I型调节T细胞:小鼠和男性
定义TR1细胞的免疫系统已经发展出一种调节免疫功能的机制,并确保炎症反应不会升级远远超出对宿主受益的升高。I型调节t(TR1)细胞(CD4 + T细胞的子集)被认为是通过抑制髓样细胞和其他T细胞的炎症程序来促进免疫反应的重要作用(1)。tr1细胞被构成了T细胞的调节子集,但与常规Treg不同,因为它们不会组成性地表达FOXP3(1)。TR1细胞已与多种临床相关疾病的结果有关。在接受造血干细胞移植并表现出同种异体移植的SCID患者中,他们发现它们相对较为焦点(2)。这些TR1细胞主要是供体的起源和缓解的移植物 - 主宿主(GVHD),通过诱导抗原特异性
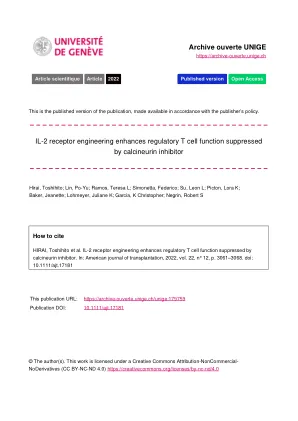
HIRAI, Toshihito 等人。IL-2 受体工程增强了受
图 1 Ortho IL-2 和他克莫司协同增加 %ortho Tregs。用 ortho IL-2R β (ortho Tregs) 转导的 Foxp3 GFP + Tregs 和幼稚 Tcons 与 CD3/CD28 珠一起孵育。按指示添加 Wt IL-2 (1000 IU/ml)、ortho IL-2 (100 000 IU/ml) 和他克莫司 (100 ng/ml),每 2 天补充一次。(A) 共培养细胞的代表性伪彩色图。(B-D) 箱线图显示第 4 天的 Tcons (B)、ortho Tregs (C) 和 ortho Tregs 在总细胞中的比例 (D)。在 2 个独立实验中的 1 个代表性实验中对三个重复孔进行量化。 * < 0.05, ** < 0.01, *** < 0.001;p 值由 Dunnett 检验计算得出,将他克莫司 (−) 组或他克莫司 ( + ) 组中的每列与 PBS 对照 (灰色条) 进行比较。ns,不显著。

羊水干细胞衍生的细胞外囊泡教育2型常规树突状细胞以在多发性硬化症小鼠模型中营救自身免疫性疾病
摘要树突状细胞(DC)是免疫反应的必不可少的编排,代表了自身免疫性疾病中免疫调节的潜在靶标。人类羊水的分泌组在免疫调节因素中很丰富,细胞外囊泡(EV)是重要的组成部分。然而,这些电动汽车对树突状细胞子集的影响仍未得到探索。在这项研究中,我们研究了高度纯化的树突状细胞亚群和源自羊水干细胞系(HAFSC-EVS)的EV之间的相互作用。我们的结果表明,通过CD29受体介导的内在化的常规树突状细胞2(CDC2)优先吸收了HAFSC-EV,从而导致耐受性DC表型,其特征在于表达降低和产生炎性介体的产生。此外,与媒介物处理的对照细胞相比,在共培养系统中使用HAFSC-EV处理Cdc2细胞会导致表达调节性T细胞标记FOXP3的T细胞比例更高。此外,将HAFSC EV处理的Cdc2s转移到EAE鼠标

1胰岛素B肽-MHC II类特异性嵌合抗原...
引言自身免疫1型糖尿病(T1D)是由T细胞介导的胰腺中Langer-Hans胰岛胰岛产生的β细胞破坏引起的。Treg是抑制CD4 + T细胞,通常起作用以限制自动反应效应的T细胞响应并预防自身免疫性(1-3)。Tregs具有异常的细胞因子和基因表达谱并减少了抑制性功能,导致概念是恢复Treg功能的策略可能是治疗或预防自身免疫性的有希望的方法(4-8)。使用T1D动物模型的大量研究表明,Treg的治疗恢复可以预防疾病的疾病。临床研究表明,这种方法在人类中的安全性,但到目前为止,功效的证据受到限制(9-11)。的理由是,迄今为止,T1D中Treg治疗的所有临床试验都使用了多克隆细胞,这意味着只有一小部分注入细胞是针对疾病与疾病相关的抗原的特异性的(12)。

先天性免疫缺陷症中的转录因子缺陷
转录因子 (TF) 是调节免疫系统发育、维持和功能的关键成分。因此,某些 TF 的单基因缺陷会导致先天性免疫缺陷 (IEI),其临床意义深远,包括感染、恶性肿瘤,在某些情况下还会导致严重的过敏性炎症。本综述探讨了以严重过敏性为定义临床表型的 IEI 背后的 TF 缺陷,包括 STAT3 功能丧失、STAT6 功能获得、FOXP3 缺陷和 T-bet 缺陷。这些疾病为了解过敏性炎症的病理生理学提供了宝贵的见解,拓展了我们对罕见单基因和常见多基因过敏性疾病的理解。基因检测的进展可能会发现与过敏性相关的新 IEI,丰富我们对过敏性炎症分子通路的理解。单基因疾病的识别对患者的预后、治疗计划和遗传咨询有深远的影响。因此,对于患有严重、早发性过敏症的患者,考虑 IEI 至关重要。本综述强调需要继续研究 TF 缺陷,以加强我们对过敏性疾病的理解和管理。
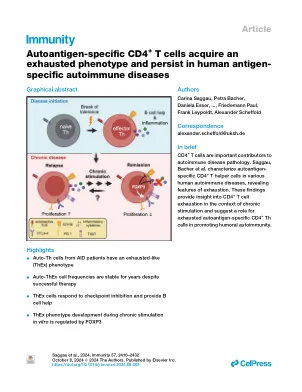
自动抗原特异性CD4 + T细胞获得了疲惫的表型,并持续存在于人类抗原特异性自身免疫性疾病
促炎性自身抗原特异性CD4 + T助手(自动三)细胞是自身免疫性疾病(AIDS)的中央编排者。我们的目的是通过结合人类白细胞抗原(HLA) - 四聚体基于定义的自身抗原在人类辅助中表征这些细胞,基于基于激活的多维外体离体分析。在Aquaporin4-抗体阳性神经瘤谱谱障碍(AQP4-NMOSD)患者中,自动三细胞表达CD154,但增殖能力和促炎性细胞因子大大降低。相反,与耗尽相关的共抑制受体与Foxp3一起表达,即规范调节T细胞(Treg)转录因子。自动三细胞在体外对检查点抑制作用并提供有效的B细胞帮助。细胞。最初在癌症和慢性感染中描述了,我们的数据表明T细胞耗尽是对跨辅助类型的慢性(自我)刺激的常见机制,并将示例性的CD4 + T细胞与体液自身免疫反应联系起来,这对治疗靶向的意义。

可溶性 RAGE 可预防 1 型糖尿病扩增功能性调节性 T 细胞
1 型糖尿病是一种无法治愈的自身免疫性疾病,由于可重复性危机,有希望的治疗方法的临床转化受到阻碍。在这里,两个独立的研究中心通过短期施用晚期糖基化终产物受体 (sRAGE) 拮抗剂来预防小鼠糖尿病。用 sRAGE 治疗可增加胰岛、胰腺淋巴结和脾脏内的调节性 T 细胞 (T regs),从而提高胰岛胰岛素的表达和功能。T reg 耗竭可消除糖尿病保护作用,并显示依赖于使用基因敲除小鼠拮抗 RAGE。用 RAGE 配体治疗的人类 T regs 下调了抑制、迁移和 T reg 稳态的基因 (FOXP3、IL7R、TIGIT、JAK1、STAT3、STAT5b、CCR4)。 sRAGE 逆转了抑制功能的丧失,其中 T regs 增加了增殖并抑制了常规 T 细胞分裂,证实了 sRAGE 扩增了功能性人类 T regs。这些结果突出了 sRAGE 是一种预防糖尿病的有吸引力的治疗方法,在多个研究中心和人类 T 细胞中显示出有效性和可重复性。