XiaoMi-AI文件搜索系统
World File Search SystemGFP

基于CAS12A的基因编辑系统,用于疫霉菌Infestans揭示了诱发仪的单相表达
fi g u r e 1在疫霉菌中核酸内切酶的表达表达。(a)五个代表性菌株的免疫印迹,用编码编码绿色荧光蛋白(GFP)标记的核酸酶的催化型核酸酶,PSNLS-DCAS9-GFP的质粒转化,用抗GFP探测。nc1和nc2是阴性对照,即分别表达另一种蛋白质和未转化的1306的菌株。蛋白质的预期大小为194 kDa。(b)表达PSNLS-DCAS9-GFP的转化剂的荧光显微照片,显示蛋白质在菌丝内的核的定位。GFP,明亮的场和合并的通道被上下显示,比例尺等于10 µm。 (c)用抗Cas12a探测的质粒转化的菌株的免疫印迹。nc是未转化的祖细胞菌株。从图像中删除了T9和NC之间的一个空车道。蛋白质的预期大小为153 kDa。(d)表达PSNLS-CAS12A-GFP的转化剂的共聚焦图像,显示左侧的菌丝,右侧显示孢子囊。GFP,明亮的场和合并的通道被上下显示,比例尺等于10 µm

原位缺失揭示了拟南芥中细胞内免疫受体基因及其共表达基因表达的调控成分
图 8 LHY1 和 bHLH28 在 SNC1 表达调控中的作用。(a)来自 DAP-seq 数据库的 SNC1 基因座中两个转录因子 LHY1 和 bHLH28 的结合。这是从浏览器图像中重新绘制的。结合用彩色块表示,高度代表检测到的结合水平。(b)野生型或 bon1 中突变体 bhlh28 和 lhy1 的生长表型。植物在 22°C 下 16 小时/8 小时光照下生长。(c)通过定量实时聚合酶链反应 (qRT-PCR) 测定 bhlh28 和 lhy1 单突变体和双突变体与 bon1 的相对 SNC1 表达。肌动蛋白被用作参考基因,并将表达水平与 Col-0 进行比较。显示的是三次生物学重复的平均值,误差线表示标准差。不同字母表示基因型之间的统计学显著差异(p < 0.05,学生 t 检验)。(d)LHY1-GFP 和 bHLH28-GFP 与 SNC1 启动子区的染色质免疫沉淀 (ChIP)-qPCR 分析。分别在“A”位点和“B”位点(如 a 所示)检测到 LHY1-GFP 和 bHLH28-GFP 的结合。显示了两个独立生物学重复的数据。“N”位点(如 a 所示)是 SNC1 基因体上的一个区域,在 DAP-seq 数据库中未检测到 LHY1 或 bHLH28 的结合信号。“GFP”是用抗 GFP 抗体孵育的样品,“NoAb”是不含抗 GFP 抗体的样品。不同字母表示通过单因素方差分析(ANOVA)得出的基因型间统计学差异具有显著性(p < 0.05)[彩色图可在 wileyonlinelibrary.com 上查看]

荧光染料标记改变了...
摘要:荧光染料标记是分析活生物体中纳米颗粒的命运的常见策略。然而,在多大程度上可以改变原始纳米颗粒生物分布的程度。在这项工作中,两种广泛使用的荧光染料分子,即Atto488(Atto)和Sulfo -Cy5(S -CY5),已共价附加到一个良好的CXCR4靶化的自我组合蛋白Nananoparticle(已知T222 -GFP -gfp -h6)上。随后已经将标记为T22 -GFP -H6 -ATTO和T22 -GFP -H6 -S -CY5纳米颗粒的生物分布与不同的CXCR4+肿瘤小鼠模型中的非标签纳米粒子的生物分布进行了比较。我们观察到,虽然父母T22 -GFP -H6纳米粒子主要是在CXCR4+肿瘤细胞中积累的,但标记为T22 -GFP -H6 -ATTO和T22 -GFP -H6 -SCY5纳米粒子在非生物分配模式中表现出急剧变化,累积的含量是巨大的,累积了,累积了,累积了,累积了,累积了,累积了。肿瘤靶向能力。因此,在靶向纳米级药物输送系统的设计和开发过程中,应在目标和非目标组织摄取研究中避免使用此类标记分子,因为它们对纳米材料的命运的影响可能会导致实际的纳米粒子生物分布的遗迹。

DNA使用Omni Bead Ruptor 12珠磨机均质器从大麻sativa中提取DNA,以进行样品制备。
图4:a)MDA-MB-231细胞被绿色 - 肾上腺胶体易感病毒感染,并使用紫霉素选择了稳定的转导细胞。每周一次通过IVIS系统确定生物发光信号的强度六周。b)用红色葡萄糖和GFP标记的双重标记的MDA-MB-231细胞。可以通过两个FACS分析检测GFP报告基因表达。c)使用Nuance多光谱成像系统测量HT1080细胞中的GFP表达。

利用新型天然产物抑制剂阿扎洛霉素 F4a 靶向自噬肽酶 ATG4B 治疗晚期胃癌
构建自噬通量分析模型,以分析我们之前报道的自噬通量变化(14)。简而言之,酸性溶酶体环境导致 pH 敏感的绿色荧光(GFP)减少,红色荧光(RFP)保持,自噬体同时显示 mRFP 和 GFP 信号,而溶酶体显示高 mRFP 信号和低 GFP 信号,这可用于指示自噬体与溶酶体的融合步骤。正如预期的那样,雷帕霉素(Rap,一种自噬诱导剂)处理导致 GCa 细胞中出现更多的红色荧光斑点,而 Am-F4a 和 Bafilomycin(Baf,一种自噬抑制剂)处理的细胞显示出高水平的黄色斑点,来自 GFP 和 RFP 的混合物(图 3b)。结果表明,Am-F4a 可能有效抑制自噬通量。为了检验 Am-F4a 对自噬的影响是否由溶酶体功能障碍引起,用 Lyso-

最大化吞吐量的定量西方结果
(a)上面显示的是来自单个LEO运行的数据。使用重组GFP蛋白的8点标准曲线在弹药筒上三次。墨盒2、3和4用于定量来自2种不同批次和各种剂量的转导细胞裂解物中的GFP表达。重组蛋白以2.5 ng/ml的速度作为校准器在所有墨盒上作为用于墨盒校正的校准器。(b)上面显示的是来自单个JES运行的数据。使用重组GFP蛋白的8点标准曲线运行,并用于定量来自2个不同批次转导的细胞裂解物中的GFP表达。总体而言,每杰西斯运行的毛细血管数量有限,只能容纳标准曲线的一个复制和有限数量的样品。在杰西系统上处理96个样本至少需要4次运行和12小时。

斑马鱼药物筛选证实厄洛替尼是 T 细胞急性淋巴细胞白血病中 Wnt/β-Catenin 信号和自我更新的抑制剂
图 1:6xTCF/LEF-miniP:GFP 斑马鱼系对 Wnt 信号通路的小分子调节剂产生可量化的反应。(A) Wnt/β-catenin GFP 报告基因 6xTCF/LEF-miniP:dGFP 转基因斑马鱼系的示意图。(B) 受精后 48 小时 (hpf) 的 6xTCF/LEF-miniP:dGFP 斑马鱼幼虫。GFP 荧光表明 Wnt 信号活跃,尾鳍 (虚线框) 用于量化。(C) 用 DMSO、Wnt 通路抑制剂 XAV939 或 Wnt 通路激活剂 BIO 处理 24 小时的 6xTCF/LEF- miniP:dGFP 幼虫中的代表性尾鳍荧光。从左到右的面板显示了明场图像、GFP 荧光和使用 ImageJ 软件对荧光进行标准化阈值处理。图中标出了与 DMSO 相比荧光增加或减少的百分比。比例尺 = 500 μm。

CRISPR/Cas9 介导的日本柳杉 (Cryptomeria japonica D. Don) 定向诱变
明显分为阳性和阴性;根据我们的观察,没有子叶表现出嵌合 GFP 荧光(图 4a-j)。在具有活性 GFP 的绿色发芽体细胞胚中,由于叶绿素自发荧光强,几乎观察不到 GFP 荧光;相反,在胚基部的愈伤组织中观察到 GFP 荧光(图 4d,i)。为了研究子叶体细胞胚中的嵌合性,使用 8-30 个子叶胚(来自 6 个品系的 139 个)进行了测序分析。来自品系#47-2 的一个子叶胚在一个体细胞胚中有两种修饰模式。然而,在其他品系中,突变模式在单个子叶胚中明显分开(图 4k)。接下来,通过分析 4 个品系(分别为 #42 - 2、#18、#31 - 2 和 #11)中各 10 个通过体细胞胚胎发生再生的幼苗,分析了突变模式的稳定性。

1型糖尿病的儿童和青少年的地中海饮食依从性和血糖控制介导的植物免疫
了解CNC@PDA@Zn 2+诱导的NSLTP2定位变化的功能意义,我们测试了NSLTP2的抗病毒功能。虽然NBLTP1在TMV感染中的作用得到很好的特征,但我们系统地探索了NSLTP2在TMV感染中的作用。应力表达分析表明,在2 dpi的接种叶中,NSLTP2的表达与TMV-GFP显着增加,其表达在6 dpi的全身性叶片中也显着升高(图S7),这表明NSLTP2在抗病毒防御中的潜在作用。接下来,我们利用烟草病毒(TRV) - 介导的基因沉默来分析NSLTP2在TMV抗性中的作用。随后,本尼亚氏菌叶具有TRV1 + TRV2(TRV:00)或TRV1 + TRV2:NSLTP2:NSLTP2(TRV:NSLTP2)12天,RT- qPCR分析显示,NSLTP2在TRV中的NSLTP2表达显着 5b)。 然后,我们用TMV-GFP机械地接种了第6和7叶,并观察到在2、4和6 DPI下紫外线下的GFP运动以跟踪TMV分布。 如图所示 5A,在接种叶片的接种叶片中观察到GFP荧光信号,而QPCR分析表明,沉默的植物中TMV-GFP核酸水平明显高于对照组(图5b)。然后,我们用TMV-GFP机械地接种了第6和7叶,并观察到在2、4和6 DPI下紫外线下的GFP运动以跟踪TMV分布。如图5A,在接种叶片的接种叶片中观察到GFP荧光信号,而QPCR分析表明,沉默的植物中TMV-GFP核酸水平明显高于对照组(图5C)。 在4 DPI时,GFP信号出现在沉默的植物的全身叶子中,而在控制植物的全身叶子中未检测到GFP信号。病毒核酸的QPCR分析产生了相似的结果(图 5d)。 5e)。5C)。在4 DPI时,GFP信号出现在沉默的植物的全身叶子中,而在控制植物的全身叶子中未检测到GFP信号。病毒核酸的QPCR分析产生了相似的结果(图5d)。5e)。通过6 DPI,GFP信号在沉默的植物中更为明显,TMV-GFP在其全身叶片中显着积累(图这些结果表明NSLTP2沉默促进了TMV-GFP感染。随后,我们比较了
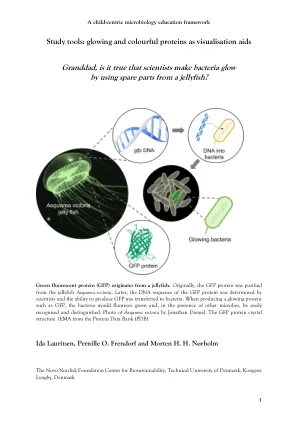
爷爷,科学家的确使细菌发光...
GFP的故事也有一个科学的“尤里卡时刻”(故事讲的是,古希腊科学家阿奇米德大喊“尤里卡!eureka!”他进入水后,使他意识到科学原则,流离失所的水量等于淹没物体的体积)。同样,在编码“绿色发光”蛋白的基因(现在称为绿色荧光蛋白)的基因编码之后,已经发现了名为GFP及其序列确定的序列,Martin Chalfie将其转移到细菌和蠕虫中,这足以使这些高度不同的生物体使这些高度不同的有机体光亮绿色 - Eureka -eureka!在接下来的几年中,罗杰·蒂恩(Roger Tsien)领导了研究工作,这些研究将大大提高GFP的特性,以使其成为通用的研究工具。例如,它们还制作了红色荧光蛋白。共同通过2008年的诺贝尔化学奖认可了这些发展,因为GFP完全改变了我们可以研究微观现象的方式。