XiaoMi-AI文件搜索系统
World File Search SystemGFP
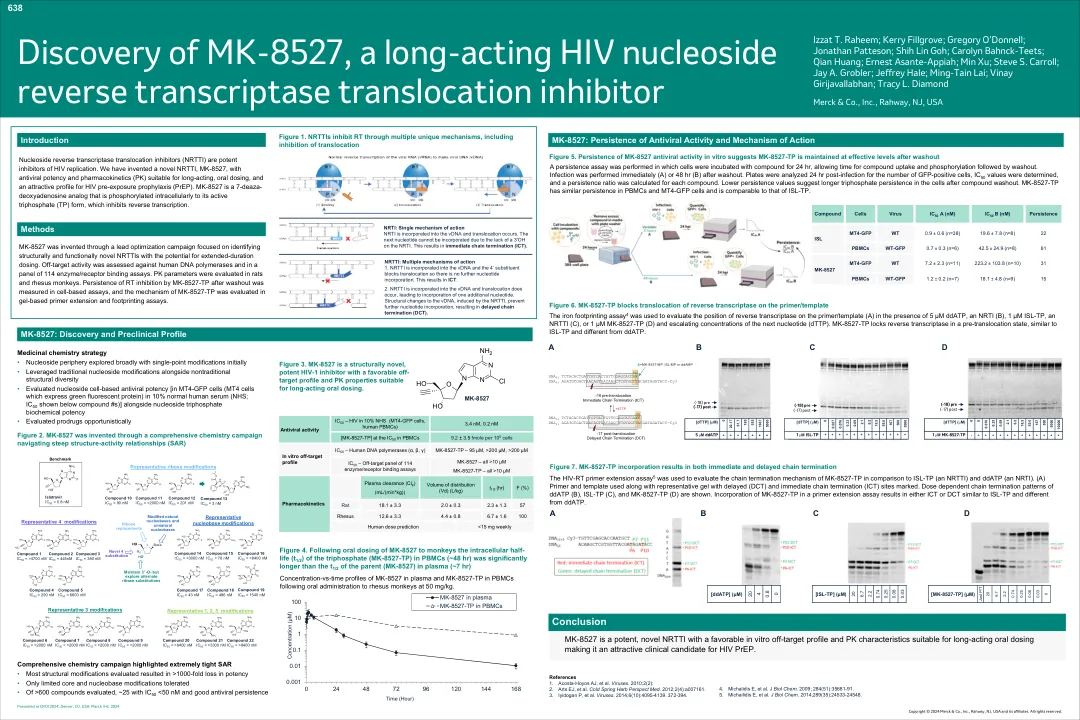
发现MK-8527,一种长效HIV HIV核苷逆转录酶易位抑制剂 对第二代集成酶抑制剂的抗性是一种罕见的... CD4靶向的mRNA tovilly tat的mRNA逆转HIV-1潜伏期 程序指南
进行了一种持久性测定,将细胞与化合物孵育24小时,从而使化合物摄取和磷酸化随后进行清洗。洗涤后立即(a)或48小时(b)立即进行感染。板对GFP阳性细胞的数量,确定IC 50值,并计算每种化合物的持续比率。较低的持久性值表明复合冲洗后细胞中三磷酸的持久性更长。MK-8527-TP在PBMC和MT4-GFP细胞中具有相似的持久性,并且与ISL-TP相当。

体外研究
乳腺癌是一种复杂的异质疾病,具有独特的分子亚型,它限制了每个亚型的优化治疗策略的发展。癌症基因疗法和晚期/难治性癌症的潜在疗法可能有望成为乳腺癌。结合了肿瘤 - 循环脂质纳米颗粒(LNP)和诱导的caspase-9(IC9)mRNA,我们旨在制定一种新型的治疗策略,以解决难治性乳腺癌。LNP的抗肿瘤作用:MDA-MB231,SKBR3和MCF-7。肿瘤细胞。通过逆转录酶定量PCR评估了与凋亡相关的基因。lnps可以有效地将包裹的GFP mRNA传递给所有三种癌细胞系(> 80%GFP表达。在目标细胞中)。此外,用IC9 mRNA(IC9-LNP)和CID封装的LNP在体外对所有癌细胞系显示了细胞毒性活性。有趣的是,在癌细胞系中,对IC9基因疗法的敏感性是良好的。IC9-LNP具有CID诱导的对SKBR3和MDA-MB231细胞的有效细胞毒性作用,但仅对MCF7细胞产生轻度的细胞毒性作用。量化相关基因的定量表明,高BAX/BCL-2比率可能与IC9-LNPÞCID易感性有关。 因此,使用IC9-LNP和CID的癌症基因治疗可能是治疗乳腺癌的有希望的替代方法,尤其是对于侵略性乳腺癌而言。 ©2022 Elsevier Inc.保留所有权利。量化相关基因的定量表明,高BAX/BCL-2比率可能与IC9-LNPÞCID易感性有关。因此,使用IC9-LNP和CID的癌症基因治疗可能是治疗乳腺癌的有希望的替代方法,尤其是对于侵略性乳腺癌而言。©2022 Elsevier Inc.保留所有权利。

蓝藻聚球藻 PCC 7002 中基因表达的光遗传学控制
光合蓝藻可在生物技术中用作环境可持续的细胞工厂,将二氧化碳转化为多种生物化学物质。然而,缺乏可用于精确和动态控制基因表达的分子工具,阻碍了代谢工程,并导致产品滴度低。光遗传学工具能够以高可调性和可逆性实现光调节的基因表达控制。到目前为止,它们在蓝藻中的应用有限,物种之间的可转移性仍不清楚。在本研究中,我们在聚球藻 PCC 7002 中表达了蓝光抑制的 YF1/FixJ 和绿/红光响应的 CcaS/CcaR 系统,并使用 GFP 荧光测定和 qRT-PCR 表征了它们的性能。非蓝藻来源的 YF1/FixJ 系统表现出较差的性能,最大动态范围为 1.5 倍,尽管采取了几个步骤来改进这一点。相比之下,源自蓝藻 Synechocystis sp. PCC 6803 的 CcaS/CcaR 系统对光波长和强度反应良好,在绿光照射 30 分钟后观察到蛋白质荧光输出增加 6 倍。监测 GFP 转录水平使我们能够量化转录激活和失活的动力学,并测试多个绿光/红光和光/暗循环对系统性能的影响。最后,我们通过对 pCpcG2 输出启动子进行有针对性的遗传修饰,提高了绿光下 CcaS/CcaR 系统的活性。本研究详细描述了 Synechococcus sp. PCC 7002 中 CcaS/CcaR 系统的行为,并强调了跨物种转移光遗传学工具的复杂性。

CALIPERS:用于表型分析实验和再生研究的细胞周期感知活体成像
1. 意大利帕维亚大学合成生理学实验室 2. 意大利米兰人类科技城 3. 意大利都灵大学“Guido Tarone”分子生物技术中心 * 通讯作者:francesco.pasqualini@unipv.it;moises.disante@unipv.it 摘要 在活细胞成像中测量细胞结构和功能以及细胞周期进程一直很有挑战性,因为荧光泛素细胞周期指示剂 (FUCCI) 和大多数表型传感器都使用绿色 (GFP) 和红色 (RFP) 荧光蛋白。我们介绍了 CALIPERS,一种用于表型分析实验和再生研究的细胞周期感知活细胞成像方法。CALIPERS 使用一种名为 FUCCIplex 的定制 FUCCI 传感器,该传感器与基于 GFP 和 RFP 的传感器进行光谱多路复用。为了证明 CALIPERS 的广泛应用范围,我们用上皮和人类诱导性多能干细胞多色报告基因系在增殖、迁移、心脏药物检测和再生医学研究中对其进行了验证。正文组学和成像技术的融合为基础科学 1,2 、药物检测 3 和再生医学 4 中的细胞表型的高级评估提供了动力。此外,参考人类诱导性多能干细胞 (hiPSC) 和多谱系分化的强大协议(例如心肌细胞、hiPSC-CM)增强了可重复性 5 ,并将表型分析工作扩展到类器官 6,7 和器官芯片 8,9。然而,细胞周期 (CC) 可能会混淆这些研究,因为随着细胞在分裂后生长(G1 期)、复制其 DNA(S)、在随后的分裂前生长(G2)或分裂(M)10 ,基因表达、形态和行为会发生变化。这在分子表型分析中得到了很好的解决,因为由于同时测量许多 CC 基因/蛋白质 11 ,大多数组学研究都具有 CC 感知能力。然而,基于成像的 CC 感知表型分析具有挑战性。通过对 G1/S/G2/M 标记物进行特定染色,可以使化学固定样品的结构表型分析具有 CC 感知能力 12 。然而,功能表型分析只有通过活细胞成像 13,14 才有可能,目前很难使用标准荧光显微镜同时评估 CC 以及细胞结构和功能。事实上,绿色和红色荧光蛋白 (GFP、RFP) 为荧光泛素细胞周期指标 (FUCCI) 10 和大多数表型传感器 15 提供动力。在这里,我们引入了一个可复用的 FUCCI 传感器 FUCCIplex,并展示了 CC 感知实时成像,用于人类上皮细胞(HaCaT,图 1)和 hiPSC(图 2)中的表型分析实验和再生研究(CALIPERS)。为了创建 FUCCIplex,我们将 fastFUCCI 传感器 16 中的 GFP 和 RFP 替换为 miRFP670(iRFP)和 mTurquoise2(CFP)。因此,FUCCIplex 细胞的细胞核在 G1 中包含 CFP,在 G1-S 过渡期包含 CPF 和 iRFP,在 S/G2/M 期仅包含 iRFP(图 1a)。为了展示 CALIPERS,我们在 HaCaT 细胞中共表达了 FUCCIplex 和肌动蛋白结合肽 RFP-LifeAct 17,并使用 40 小时以上的活细胞荧光成像来追踪细胞在每个 CC 期所花费的时间(图 1b 和补充视频 1 和 2)。我们证实 HaCaT 细胞约 40% 的时间处于 G1 期,其余时间处于 S/G2/M 期,这与使用 FUCCIplex 或 DNA 标记在静态图像和流式细胞术实验中测得的 CC 期占有率一致(图 1c-d 和扩展图 1)。此外,我们开发了一个开源插件,可将 CFP 和 iRFP 强度转换为 FUCCIphase 信号,该信号可追踪 CC 完成百分比并实现 CC 感知的形态和运动分析(图 1e、补充视频 3 和扩展图 2)。


协会9(CRISPR/CAS9)最新技术编辑gen ...
糖尿病(DM)是一种代谢疾病,由胰腺β细胞的自身免疫反应引起的1型DM,以及由于人体无法最佳使用胰岛素而引起的第二类DM。在2010年,有113,000名儿童患有1型DM,预计将增加18,000名儿童。1型糖尿病是一种疾病,需要在其治疗中进行基因编辑过程。crispr/cas9已知可以有效改变HPSCS基因(茎祖细胞基因)DM 1型。crispr/cas9在细菌和芳香族中发现,在适应性免疫中起作用。修复胰腺细胞的过程有助于人体的胰岛素调节,同时治疗DM 1型疾病本综述文献中使用的方法是对16本期刊和2本书组成的文章的评论,并通过Google Scholar,PubMed和Mendeley获得。。各种基因编辑方法,其中一种是CRISPR/CAS9和人才。有3种类型的祖细胞(HPSCS细胞)胰腺,已知能够产生胰岛素的胰腺是NKX6.1-GFP HUES8细胞,H1细胞和诺科唑细胞。在CRISPR/CAS9上,GFP的存在显示了NHEJ捐赠者存在的DSB的改进结果。GFP通过体验增加47.6%,在胰腺祖细胞(NKX6.1-GFP HUES8细胞)的推导中起作用。显示细胞增加17.77%。最后,在诺科唑细胞中,与编辑talen方法相比,已知有54.64%的分化。,CRISPR/CAS9可以是一种有效的基因编辑方法,尤其是在1型DM治疗中

一种用于裂殖酵母的快速有效的 CRISPR/Cas9 方法[...
摘要 CRISPR/Cas9 系统可实现无疤痕、无标记的基因组编辑。目前,用于裂殖酵母 Schizosaccharomyces pombe 的 CRISPR/Cas9 系统依赖于繁琐且耗时的克隆程序,将特定的 sgRNA 靶序列引入 Cas9 表达质粒中。此外,据报道,当从强 adh1 启动子持续过表达 Cas9 核酸内切酶时,它会对裂殖酵母产生毒性。为了克服这些问题,我们开发了一种改进的系统 SpEDIT,它使用从中等强度 adh15 启动子表达的针对 S. pombe 进行密码子优化的合成 Cas9 序列。SpEDIT 系统表现出灵活的模块化设计,其中 sgRNA 与自切割丁型肝炎病毒 (HDV) 核酶的 3' 端融合,从而允许 tRNA 基因序列中的 RNA 聚合酶 III 驱动 sgRNA 盒的表达。最后,在 GFP 占位符两侧加入 Bsa I 型 IIS 限制酶位点,可实现 Golden Gate 介导的一步式 GFP 替换和合成的 sgRNA 表达。SpEDIT 系统通过转化异步培养细胞,在 ade6 + 或 ura4 + 基因中生成靶点突变体,可实现 100% 的诱变效率。SpEDIT 还允许以最小的努力获得插入、标记和删除事件。还可以轻松实现两个独立非同源基因位点的同时编辑。重要的是,与目前可用的 S. pombe 编辑系统相比,SpEDIT 系统显示出更低的毒性。因此,SpEDIT 提供了一种有效且用户友好的 CRISPR/Cas9 程序,可显著改善裂殖酵母的基因组编辑工具箱。

全基因组 CRISPR 筛选确定 TLR3 信号通路的调控因子
摘要 TLR 的一个子集专门通过对内体进行核酸检测来检测进入的病原体。其中,TLR3 感知内体中双链 RNA 的异常存在,并通过激活 NF- j B 和 IRF3 启动强大的先天免疫反应。然而,控制 TLR3 调节的机制仍然不甚明了。为了确定参与 TLR3 通路的新分子参与者,我们使用 CRISPR/Cas9 技术进行了全基因组筛选。我们生成了携带 NF- j B 反应启动子的 TLR3 + 报告细胞,该启动子控制 GFP 表达。接下来用单向导 RNA (sgRNA) 文库转导细胞,用 poly(I:C) 进行连续刺激,并对 GFP 阴性细胞进行分类。通过深度测序估计的 sgRNA 富集确定了 TLR3 诱导的 NF-j B 激活所需的基因。在这些基因中,通过筛选确定了五个已知与 TLR3 通路密切相关的基因,包括 TLR3 本身和伴侣 UNC93B1,从而验证了我们的策略。我们进一步研究了前 40 个基因,并重点研究了转录因子芳烃受体 (AhR)。AhR 的消耗对 TLR3 反应有双重影响,消除了 IL-8 的产生并增强了 IP-10 的释放。此外,在暴露于 poly(I:C) 的原代人巨噬细胞中,AhR 激活增强了 IL-8 并减少了 IP-10 的释放。总体而言,这些结果表明 AhR 在 TLR3 细胞先天免疫反应中发挥作用。
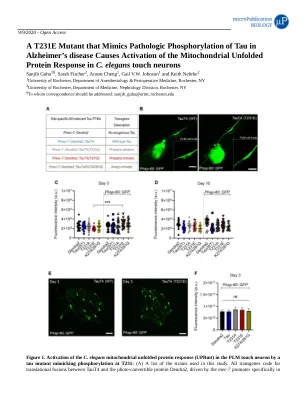
T231E突变体,在阿尔茨海默氏病中模仿Tau的病理磷酸化会导致线粒体展开的蛋白质反应激活I
触摸神经元。CRISPR-CAS9基因编辑用于将磷酸化T231A,磷酸化模拟T231E和乙酰基模拟的K274/281Q突变引入Tain4 Orf。为简单起见,这些突变体将称为T231A,T231E和K274/281Q。(b,c)第3天的触摸神经元的荧光图像,表达dendra2 :: Taut4转化融合和T231E突变体的单拷贝转基因编码。虚拟的圆圈表示PLM细胞体的位置,显示在插图中。比例尺,0.5 µm。注意,斑点荧光来自后肠中标记为GFP的HSP-60表达式。(c,d)成年第3和第10天,对面板A中列出的菌株的PLM细胞体荧光定量。数据是来自两个独立技术重复的平均值±SD。各个数据点从单独动物的单个PLM细胞中划分值(n = 25±5)。统计分析是通过Tukey的事后测试进行的双向方差分析,在比较包围样品时,*** p <0.001。请注意,左侧条形柱是指单独携带Dendra2报告基因的转基因菌株的荧光定量,而右侧则是指携带Dendra2和HSP-60记者的菌株。(e)表达整合的UPR MT报告基因P HSP-60 :: GFP和单拷贝MOSSCI插入的转基因蠕虫的代表性荧光图像。比例尺,0.5毫米。数据是平均±SD(来自两个独立生物学重复的20只动物)。(f)从面板中列出的菌株的后肠道区域中荧光信号强度定量。ns表示不显着,如通过单向方差分析计算,然后进行Tukey的多重比较测试。

速度编辑
摘要 CRISPR/Cas9 系统可实现无疤痕、无标记的基因组编辑。目前,用于裂殖酵母 Schizosaccharomyces pombe 的 CRISPR/Cas9 系统依赖于繁琐且耗时的克隆程序,将特定的 sgRNA 靶序列引入 Cas9 表达质粒中。此外,据报道,当从强 adh1 启动子持续过表达 Cas9 核酸内切酶时,它会对裂殖酵母产生毒性。为了克服这些问题,我们开发了一种改进的系统 SpEDIT,它使用从中等强度 adh15 启动子表达的针对 S. pombe 进行密码子优化的合成 Cas9 序列。SpEDIT 系统表现出灵活的模块化设计,其中 sgRNA 与自切割丁型肝炎病毒 (HDV) 核酶的 3' 端融合,从而允许 tRNA 基因序列中的 RNA 聚合酶 III 驱动 sgRNA 盒的表达。最后,在 GFP 占位符两侧加入 Bsa I 型 IIS 限制酶位点,可实现 Golden Gate 介导的一步式 GFP 替换和合成的 sgRNA 表达。SpEDIT 系统通过转化异步培养细胞,在 ade6 + 或 ura4 + 基因中生成靶点突变体,可实现 100% 的诱变效率。SpEDIT 还允许以最小的努力获得插入、标记和删除事件。还可以轻松实现两个独立非同源基因位点的同时编辑。重要的是,与目前可用的 S. pombe 编辑系统相比,SpEDIT 系统显示出更低的毒性。因此,SpEDIT 提供了一种有效且用户友好的 CRISPR/Cas9 程序,可显著改善裂殖酵母的基因组编辑工具箱。