XiaoMi-AI文件搜索系统
World File Search SystemGTPase

重组人类免疫相关 GTPase 家族 M 蛋白 (IRGM)
复原 我们建议在打开前先短暂离心此小瓶,使内容物沉至底部。请使用去离子无菌水复原蛋白质至浓度为 0.1-1.0 mg/mL。我们建议添加 5-50% 甘油(最终浓度)并分装以在 -20°C/-80°C 下长期储存。我们默认的甘油最终浓度为 50%。客户可以将其作为参考。

重组人类免疫相关 GTPase 家族 M 蛋白 (IRGM)
复原 我们建议在打开前先短暂离心此小瓶,使内容物沉至底部。请使用去离子无菌水复原蛋白质至浓度为 0.1-1.0 mg/mL。我们建议添加 5-50% 甘油(最终浓度)并分装以在 -20°C/-80°C 下长期储存。我们默认的甘油最终浓度为 50%。客户可以将其作为参考。

干扰素诱导的GTPase MXB促进疱疹病毒的capsID拆卸和基因组释放
1德国汉诺威汉诺威医学院病毒学研究所; 2抵抗 - 卓越群,德国汉诺威汉诺威医学院; 3德国慕尼黑慕尼黑技术大学病毒学研究所; 4德国弗莱堡大学弗莱堡大学医学中心病毒学研究所; 5美国普林斯顿大学普林斯顿大学分子生物学系; 6 MRC人类免疫学部门,MRC Weatherall分子医学研究所,Radcliffe医学系,牛津大学,牛津大学,英国; 7德国汉诺威 - 布劳恩斯乔格合作伙伴网站德国感染研究中心(DZIF),德国汉诺威; 8德国汉诺威汉诺威医学院的研究核心单位激光显微镜; 9德国慕尼黑的德国感染研究中心(DZIF),德国慕尼黑1德国汉诺威汉诺威医学院病毒学研究所; 2抵抗 - 卓越群,德国汉诺威汉诺威医学院; 3德国慕尼黑慕尼黑技术大学病毒学研究所; 4德国弗莱堡大学弗莱堡大学医学中心病毒学研究所; 5美国普林斯顿大学普林斯顿大学分子生物学系; 6 MRC人类免疫学部门,MRC Weatherall分子医学研究所,Radcliffe医学系,牛津大学,牛津大学,英国; 7德国汉诺威 - 布劳恩斯乔格合作伙伴网站德国感染研究中心(DZIF),德国汉诺威; 8德国汉诺威汉诺威医学院的研究核心单位激光显微镜; 9德国慕尼黑的德国感染研究中心(DZIF),德国慕尼黑

通过工程模式识别受体增强植物的广谱抗性
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本版的版权持有人于2025年3月7日发布。 https://doi.org/10.1101/2025.03.02.641080 doi:Biorxiv Preprint

鉴定针对 GTPase 的潜在抑制剂——...
背景:K-Ras 基因突变是各种癌症中最常见的基因变异之一,抑制 RAS 信号传导在治疗实体瘤方面显示出良好的效果。然而,寻找能与 RAS 蛋白结合的有效药物仍然具有挑战性。这促使我们探索可以抑制肿瘤生长的新化合物,特别是对于携带 K-Ras 突变的癌症。方法:我们的研究使用生物信息学技术,如 E-药效团虚拟筛选、分子模拟、主成分分析 (PCA)、超精度 (XP) 对接和 ADMET 分析,以确定 K-Ras 突变体 G12C 和 G12D 的潜在抑制剂。结果:在我们的研究中,我们发现阿法替尼、奥希替尼和羟氯喹等抑制剂对 G12C 突变体有强烈的抑制作用。同样,羟嗪、珠氯噻嗪、氟奋乃静和多沙普仑是 G12D 突变体的有效抑制剂。值得注意的是,这六种分子都对突变结构中存在的 H95 隐蔽沟表现出高结合亲和力。这些分子在分子水平上表现出独特的亲和机制,疏水相互作用进一步增强了这种亲和机制。分子模拟和 PCA 揭示了在开关区域 I 和 II 内形成了稳定的复合物。这在三种复合物中尤为明显:G12C-奥希替尼、G12D-氟奋乃静和 G12D-珠氯哌噻吨。尽管 K-Ras 中的开关 I 和 II 具有动态特性,但抑制剂的相互作用保持稳定。根据 QikProp 结果,与 sotorasib 相比,所选分子的性质和描述符在可接受范围内。结论:我们成功地鉴定了 K-Ras 蛋白的潜在抑制剂,为开发针对 K-Ras 突变驱动的癌症的靶向疗法奠定了基础。

免疫连接基因通过与高尔基功能和ARF-1 GTPase相关的膜应激途径刺激
感染反应和其他免疫相关基因(ILG)首先在秀丽隐杆线虫中命名 - 基于病原体挑战的表达,但是当脂质代谢受到干扰时,许多人也会上调。为什么病原体攻击和代谢变化两个增加ILGS尚不清楚。我们发现,当秀丽隐杆线虫中分泌细胞器的膜膜的磷脂酰胆碱(PC)水平变化时,ILG被激活。RNAi靶向ADP-核糖基化因子ARF-1(破坏高尔基体和分泌功能)也激活了ILGS。低PC限制ARF-1功能,这表明通过脂质代谢进行ILG激活的机制,这是作用于ER外的膜应激反应的一部分。RNAi在两个GFP替代者的分泌中发现了缺陷,并积累了病原体响应的补体C1R/C1S,UEGF,BMP1(CUB)域融合蛋白。我们的数据认为,某些ILG的上调是对贩运变化的协调反应,并且可能采取行动来抵消对分泌功能的压力。

鉴定在上皮卵巢癌中具有治疗价值的新型RAN GTPase的抑制剂
Singh Singh 1,Catine E. Greeson 2,Misy Fang 3,Yasmin N. Laymon 4,Vishai Khivansara 3,Yavin 3,Yavu T. Dura 1,荷兰人4,荷兰人4,Yixiang li 1,Musannad Abu-Remaleh 1,Abu-Remaleh 1,xinah li 1,xinah li 1,xinah li 1,xinah li 1,xinah li 1,xinah li s.SOL 2,Fraene Frama-Walton 2,Luis Hernandaz 2,Miguel P Balldoero 2,Bryn M. Leant Mean Mean Evely 2,Iolda Venndrell 8,Roman Fiker 8,Kessilla 8,Kessilla 8,Preparla C. Cukhale,Signoretti先知4,Peblo D. Sepolp 2,Earp 2,Earp D. Sepolp D.Sepolp D.

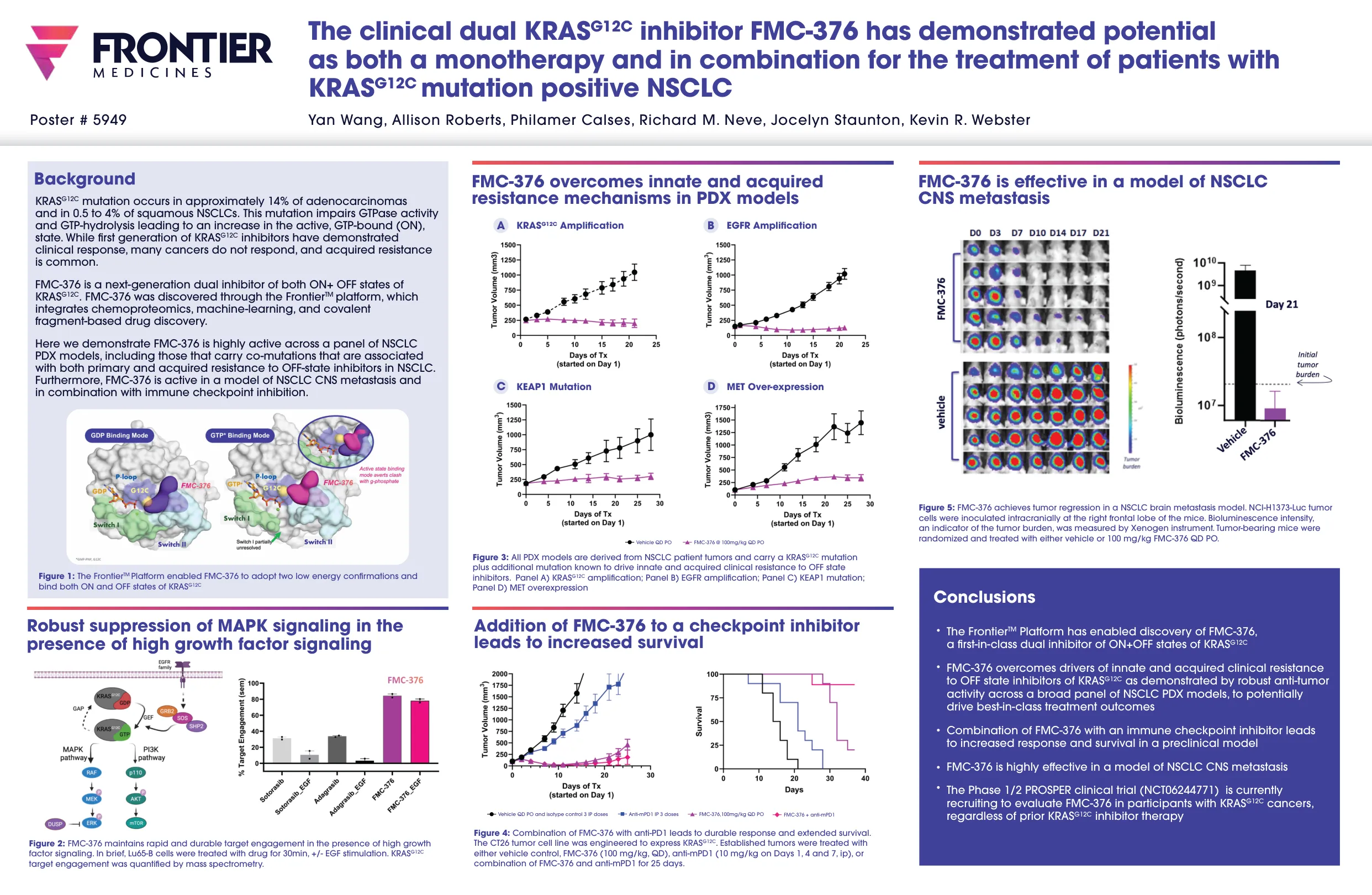
临床双KRASG12C抑制剂FMC-376 ...
KRAS G12C突变发生在大约14%的腺癌中,在0.5至4%的鳞状NSCLC中发生。该突变会损害GTPase活性和GTP - 溶解度,从而导致活性,GTP结合(ON)状态增加。虽然第一代KRAS G12C抑制剂表现出临床反应,但许多癌症没有反应,并且获得的抗性很常见。

RHOGDI2在免疫和癌症中的双重功能
Rho GTPases是RAS超家族的高度保守成员,最著名的是组织肌动蛋白和微管细胞骨架,从而定义细胞的形状和迁移。他们还控制着各种各样的信号通路,这些信号通路可以调节关键的生物学过程,例如囊泡转运,细胞分裂和基因转录[1-3]。Rho GTPases循环在活动的GTP结合形式和不活动的GDP结合形式之间。该活性受三类蛋白质的调节:鸟嘌呤核苷酸交换因子(GEFS)催化GDP的GDP交换为GTP激活GTPase;而GTPase激活蛋白(GAP)会增加GTPase的内在GTP水解速率并使IT失活;和鸟嘌呤核苷酸解离抑制剂(GDIS)隔离了细胞质中GDP结合的GDP结合形式,以防止GEFS或泛素介导的脱脂剂激活它们(图1)[4]。Rho GTPases及其调节剂的异常信号传导通常在许多人类癌症中发现,并且归因于几种机制[5-10]。到目前为止,已经确定了将近20个Rho GTPase家族成员的近85个Rhogefs和66个Rhogaps,其中,到目前为止,只有3个人Rhogdis已确定:rhogdi1(或Rhogdiα),Rhogdiα),Rhogdi2i2(或Rhogdiβ或rhogdiβ或ly d4-gdi)(或rhogdiβ或ly d4-gdi) γ)[8]。这三个仅驻留在细胞质中,其中rhogdi1被普遍表达[11,12]。rhoGDI2最初被认为在造血细胞中特异性表达[13,14],但随后在包括癌细胞在内的其他各种细胞类型和组织中也发现了[8]。最后,rhogdi3主要在靶向高尔基体的大脑,肺,肾脏,睾丸和胰腺中表达,并将特异性朝向RHOB和RHOG [15]。对癌症和免疫力中的RhoGDI3不太了解,因此在本综述中不会进一步讨论。rhogdi1和rhogdi2通过参与癌细胞迁移,侵袭和转移与多种人类癌症有关,因此被认为是癌症生物学的有吸引力的靶标[8]。rhogdi2由于其较低的丰富性和更限制的分布而在很大程度上保留在Rhogdi1的阴影中。但是,由于发现Rhogdi2可能在多种人类癌症和许多关键的细胞过程中起更复杂的作用,因此开始引起更多的关注。本评论重点介绍了Rhogdi1和Rhogdi2之间的相似性和差异,同时还封装了后者所显示或已提出扮演的多个角色。