XiaoMi-AI文件搜索系统
World File Search SystemH3K27me3
14-1048 技术数据表
CUTANA™ChIC/CUT&RUN 试剂盒可简化组蛋白翻译后修饰 (PTM) 和染色质相关蛋白的染色质分析。CUT&RUN 试剂盒版本 5 (v5) 现在包含额外的对照抗体 (H3K27me3)。阳性 (H3K4me3 和 H3K27me3) 和阴性 (IgG) 对照抗体与 SNAP-CUTANA™spike-in 对照配对,用于优化测定和持续测定监测 (图 2)。包含大肠杆菌 DNA 以进行数据标准化。SPRI 磁珠用于 DNA 纯化,可在整个工作流程中实现无缝多通道移液,从而最大限度地提高通量和可重复性。该试剂盒与各种输入兼容,包括源自天然、冷冻保存或交联样本的细胞或细胞核。虽然建议从 500,000 个细胞开始,但只需 5,000 个细胞即可生成可比数据。纳入对照以及与多种目标类型、样本输入和低细胞数量的兼容性,使该试剂盒成为染色质映射实验的首选解决方案。

CUTANA™ 止动缓冲液
CUT&RUN 方法 CUT&RUN 使用 CUTANA™ChIC/CUT&RUN 试剂盒进行,起始于 500k K562 细胞,含 0.5 µg IgG(EpiCypher 13-0042)、H3K4me3(EpiCypher 13-0060)、H3K27me3(EpiCypher 13-0055)或 0.125 µg CTCF(EpiCypher 13-2014)抗体,一式两份。使用 CUTANA™CUT&RUN 文库制备试剂盒(EpiCypher 14-1001/14-1002)以 5 ng DNA(或回收总量,如果少于 5 ng)进行文库制备。文库在 Illumina NextSeq2000 上运行,采用双端测序(2x50 bp)。样本测序深度为 5.5/18.8 百万个读数 (IgG Rep 1/Rep 2)、14.2/17.0 百万个读数 (H3K4me3 Rep 1/Rep 2)、24.7/18.1 百万个读数 (H3K27me3 Rep 1/Rep 2) 和 8.6/5.5 百万个读数 (CTCF Rep 1/Rep 2)。使用 Bowtie2 将数据与 T2T-CHM13v2.0 基因组比对。过滤数据以删除重复、多比对读数和 ENCODE DAC 排除列表区域。

GSK126
GSK126 是 zest 同源物 2 的组蛋白赖氨酸甲基转移酶增强子 (EZH2;IC 50 = 9.9 nM) 的抑制剂。1 它对 EZH2 的选择性优于 EZH1 (IC 50 = 680 nM) 和多种含有 SET 结构域和不含有 SET 结构域的甲基转移酶 (IC 50 s = 13->100 µM)。GSK126 (7-252 nM) 可降低表达野生型或突变型 EZH2 的多种弥漫性大 B 细胞淋巴瘤 (DLCBL) 细胞中组蛋白 H3 (H3K27me3) 上赖氨酸 27 的整体三甲基化。当以每天 50 mg/kg 的剂量给药时,它还可以减少 Pfeiffer DLBCL 小鼠异种移植模型中的肿瘤生长。 GSK126 (10 µM) 可诱导从患有 1 型糖尿病的青少年患者或无糖尿病的成年人中分离的人胰腺外分泌细胞产生胰岛素和葡萄糖刺激的胰岛素分泌。2 它还可降低 H3K27me3 水平并诱导原代人胰腺导管上皮细胞分泌胰岛素和葡萄糖刺激的胰岛素分泌。

IGF168PD124
理解染色质功能对于不清除欧洲核心中基因组调节的复杂性至关重要。染色质的基本亚基是Nu-Cleosome,它是由包裹在八个组蛋白蛋白的DNA形成的。Berger组的研究重点是研究组蛋白变体和重塑剂的进化论和功能,这是染色质调节的关键成分。在去年,Berger组证明了组蛋白变体与组蛋白的翻译后修饰至关重要,从而塑造了染色质指示转录调控。His-Tone变体H2A.X在维修DNA的机械中起着关键作用。Berger Group在拟南芥中确定了这一途径的两个关键参与者。此外,他们还表征了与调节转座活性的特定类型组蛋白变体的沉积相关的染色质重塑剂。最近该组还表明,在陆地植物的进化过程中,翻译后修饰H3K27me3将其靶标从转座子转换为控制基因沉默的顺式元素。H3K27me3的新功能有可能重塑植物的基因组。

被压抑但随时准备出发:双价性如何使发育基因保持表达状态
识别双价读取器的关键是该团队能够创建经过特殊修饰的组蛋白和核小体(DNA 以“串珠”结构缠绕在组蛋白上)。通过精心重建 DNA 和组蛋白复合物以进行定制的蛋白质相互作用分析,该团队已经证明,在双价位置,蛋白质被招募到抑制标记(H3K27me3)而不是激活标记(H3K4me3)。

发现 ORIC-944,一种具有最佳疗效的新型 PRC2 抑制剂...
注意:第 27 天 22Rv1 肿瘤切片中 H3K27me3 IHC 阳性细胞的定量图像分析。单因素方差分析,然后进行 Tukey 多重比较检验。****,p<0.0001;TGI 86%;肿瘤生长抑制 (TGI) = [1 - (TVtf - TVt0) / (TVcf - TVc0)] × 100%;用载体 (n=3) 或 ORIC-944 200 mpk QD (n=4) 处理的 22Rv1 异种移植瘤的 RNA 测序以评估来自转移性前列腺肿瘤的 87 基因多梳抑制特征 (Yu 等人,Cancer Res 2007);ORIC-944 与载体,t 检验:*,p<0.05。

使用瓜拉纳粉(Paullinia cupana)在...
口腔癌负责世界各地的许多死亡,因为它导致了由于治疗失败而导致的复发和转移。常规处理破坏了分化的肿瘤细胞,但肿瘤干细胞种群具有抗性并重新填充肿瘤。Wnt/β-catenin信号传导参与肿瘤干细胞的维持,生存,自我更新和分化及其信号传导,可以通过表观遗传修饰来调节。该项目的目的是确定控制Wnt/β-catenin信号通路及其靶标涉及的表观遗传变化,并研究道路参与肿瘤干细胞积累和口服癌细胞系的化学性。研究了三种野生口服癌菌株(Cal27 wt; SCC9 WT; SCC25 wt)和顺铂耐药性(Cal27 CISR; SCC9 CISR; SCC25 CISR)及其肿瘤干细胞群(CTT+)和非肿瘤干(CTT-temor(CTTT-))。QPCR分析,以评估基因表达和蛋白质印迹以进行蛋白质水平评估。通过细胞可行性测试确定IC50剂量的抑制剂。球体流量和鉴定的CTT+的形成细胞术。染色质免疫沉淀以识别道路的表观遗传调节。Xenoenxe检验用于研究Wnt/β-catenin途径作为治疗靶标的潜力。我们观察到表观遗传机调节基因的表达增加,例如BRD7,EZH2,KDM4C和MLL1和CTNNB1基因,该基因在抗顺铂菌株中编码β-catenin的ctNNB1基因。Wnt/β-catenin途径基因(如APC和GSK3β)在3种化学主义菌株中减少,下游FGF18和MMP7基因增加。CTT+的种群表现出参与组蛋白甲基化的基因的更大表达。β-catenin和甲基化的H3K27ME3和H3K9ME2组蛋白在顺铂抗性菌株和CTT+中也增加了。EZH2(UNC1999)和β-catenin抑制剂(ICG-001和FH535)的抑制剂降低了CTT+的群体,并降低了化学谱系中CTT+的群体,并降低了β-catenin和Ezh2蛋白。H3K27ME3用抑制剂处理后也降低了它。UNC1999治疗增加了上游APC和GSK3β基因的表达,并且对ICG-001,FH535和UNC1999的处理可有效降低CTT+中下游MMP7基因。FH535显示出降低CTT+种群的有效性,尤其是与顺铂和UNC1999结合使用时。β-catenin抑制剂单一疗法或与顺铂和UNC1999结合降低了CTT+躯干表型。在肿瘤组织中施用FH535,FH535+顺铂和UNC1999+FH535之后,肿瘤生长降低,肿瘤β-catenin,Ezh2,H3K27Me3和肿瘤干细胞标记肿瘤降低。通过化学谱系和CTT+CTT+种群中的染色质免疫沉淀,我们确定EZH2与该地区

生成CRISPR工具以研究脑器官发育过程中的表观遗传变化
1.1。真核生物中的表观遗传标记,DNA围绕组蛋白八聚体形成核小体,可以化学修饰。在组蛋白尾部进行的这些修饰,例如甲基化和乙酰化,影响染色质结构和基因可及性,而无需改变DNA序列。对这些修改对基因表达的影响需要诱导其在神经区域的收益或损失来评估因果关系。特定的修饰,H3K4ME3,与活性基因启动子相关,而H3K9ME3和H3K27ME3与转铺回归有关(Policarpi等,2022)。存在H3K4me3与转录之间的相关性,但是为了研究因果关系,需要通过组蛋白脱甲基酶诱导H3K4ME3损失的实验来确定在那里是否下调转录。
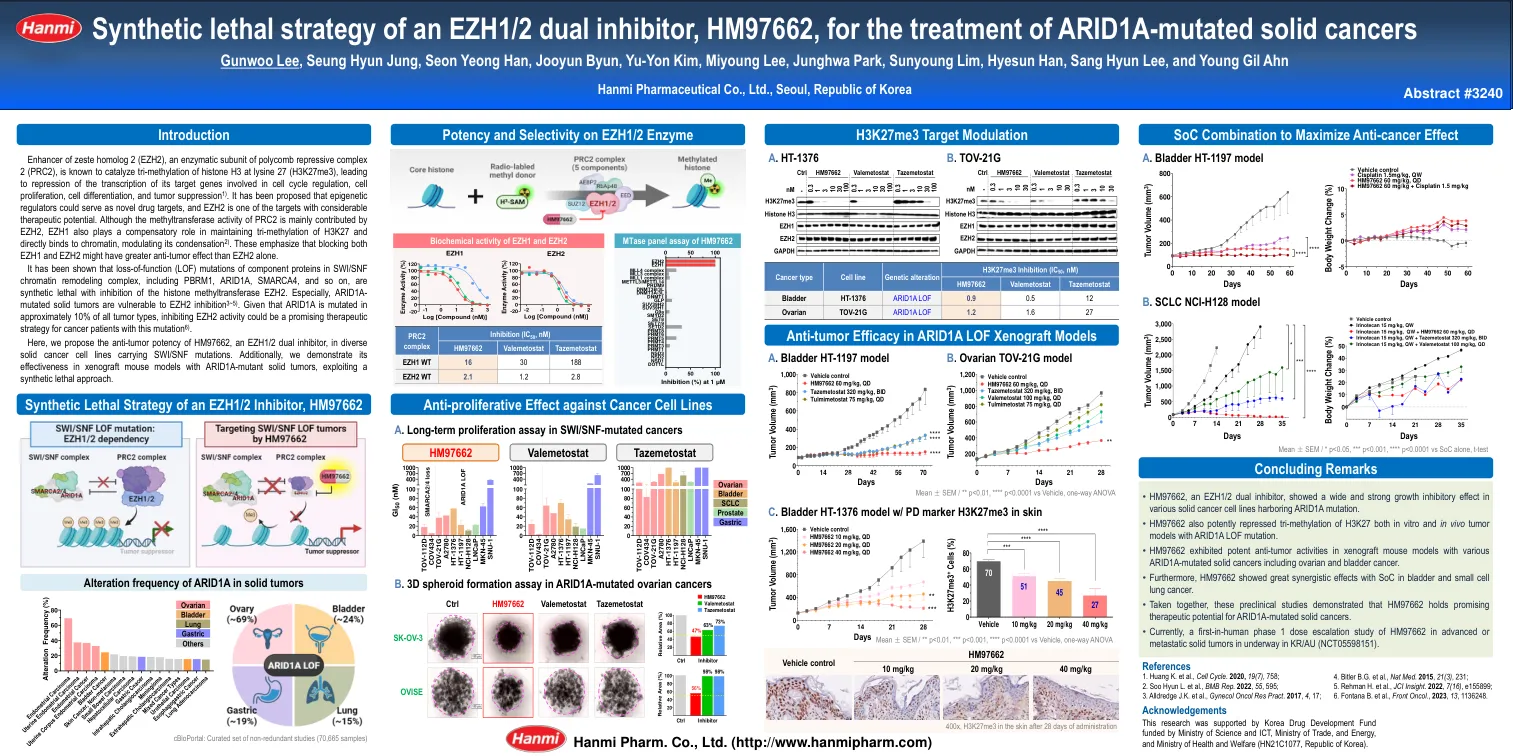
一种新颖且有效的EZH1/2双重抑制剂,HM97662表现出广泛的治疗潜力
Enhancer of zeste homolog 2 (EZH2), an enzymatic subunit of polycomb repressive complex 2 (PRC2), is known to catalyze tri-methylation of histone H3 at lysine 27 (H3K27me3), leading to repression of the transcription of its target genes involved in cell cycle regulation, cell proliferation, cell differentiation, and tumor suppression 1) .已经提出表观遗传调节剂可以用作新的药物靶标,而EZH2是具有巨大治疗潜力的靶标之一。尽管PRC2的甲基转移酶活性主要由EZH2贡献,但EZH1在维持H3K27的三甲基化方面也起着补偿性作用,并直接与染色质结合,调节其凝结2)。这些强调,与单独的EZH2相比,阻止EZH1和EZH2的抗肿瘤效应可能更大。

EZH2抑制剂联合TIGIT单克隆抗体对多发性骨髓瘤细胞的疗效研究
摘要:EZH2 是多梳抑制复合物 2 的成员之一,通过诱导下游基因组蛋白 3 位赖氨酸 27(H3K27me3)三甲基化来抑制肿瘤细胞增殖。本文表明,抑制 EZH2 后,细胞凋亡率和凋亡蛋白表达增加,而 NF- κ B 信号通路的关键分子和下游靶基因受到抑制。此外,mTOR 信号通路降低了多发性骨髓瘤 (MM) 细胞中 TIGIT 高亲和力配体 CD155 的表达。此外,EZH2 抑制剂和 TIGIT 单克隆抗体阻断剂的联合使用增强了自然杀伤细胞的抗肿瘤作用。综上所述,EZH2抑制剂不仅作为表观遗传药物发挥抗肿瘤作用,还可以通过影响NK细胞与MM细胞之间的TIGIT-CD155轴,增强TIGIT单抗的抗肿瘤作用,从而为MM患者的治疗提供新的思路和理论基础。