XiaoMi-AI文件搜索系统
World File Search SystemH3K4me3

被压抑但随时准备出发:双价性如何使发育基因保持表达状态
识别双价读取器的关键是该团队能够创建经过特殊修饰的组蛋白和核小体(DNA 以“串珠”结构缠绕在组蛋白上)。通过精心重建 DNA 和组蛋白复合物以进行定制的蛋白质相互作用分析,该团队已经证明,在双价位置,蛋白质被招募到抑制标记(H3K27me3)而不是激活标记(H3K4me3)。

WDR5-H3K4ME3表观遗传轴调节胰腺肿瘤免疫原性和免疫抑制
WDR5/MLL1-H3K4ME3表观遗传轴通常在肿瘤细胞和肿瘤浸润的免疫细胞中都被激活,以驱动肿瘤微环境中的各种细胞反应,并在造血癌中进行了广泛研究,但在肿瘤细胞和免疫细胞中,在肿瘤细胞中的相应功能仍然是肿瘤生长的肿瘤细胞。我们在这里报告说,与正常胰腺相比,WDR5在人胰腺肿瘤组织中表现出更高的表达水平。此外,WDR5表达与患者对人类结肠癌和黑色素瘤的化学疗法或免疫疗法的反应负相关。然而,在人类癌细胞中的WDR5表达与HLA水平呈正相关,并且在胰腺癌细胞的HLA-A,HLA-B和HLA-C基因的启动子区域观察到H3K4ME3富集。使用小鼠肿瘤细胞系和体内肿瘤模型,我们确定WDR5缺乏症或抑制作用在胰腺肿瘤细胞中的体外和体内抑制MHC I表达。从机械上讲,我们确定WDR5缺乏抑制MHC I(H2K1)启动子区域的H3K4ME3沉积以抑制MHC I(H2KB)转录。另一方面,WDR5耗竭导致胰腺微环境中的免疫检查点和免疫抑制细胞因子(包括TGFB和IL6)的下调。我们的数据确定WDR5不仅调节肿瘤细胞免疫原性以抑制肿瘤的生长,而且还激活了免疫抑制途径以促进肿瘤免疫逃避。在基于WDR5的表观遗传癌症的免疫疗法中应考虑选择性激活WDR5-MHC I途径和/或选择性抑制WDR5免疫检查点和WDR5-Cytokine途径。

二氢叶酸还原酶活性控制发育中的新皮质中的神经源性转变
一碳/叶酸 (1C) 代谢提供 DNA 和组蛋白甲基化所需的甲基基团,并参与维持干细胞的自我更新。二氢叶酸还原酶 (DHFR) 是 1C 代谢中的关键酶,在新皮层发育的早期阶段,在人类和小鼠神经祖细胞中高度表达。在这里,我们研究了 DHFR 在发育中的新皮层中的作用,并报告说,降低其在人类神经类器官和小鼠胚胎新皮层中的活性会加速间接神经发生,从而影响新皮层的神经元组成。此外,我们表明,降低神经祖细胞中的 DHFR 活性会导致一碳/叶酸代谢物的减少,并与 H3K4me3 水平的变化相关。我们的研究结果揭示了 DHFR 在控制新皮层发育的特定步骤中发挥的意想不到的作用,并表明 1C 代谢线索的变化会影响细胞命运转变。

间充质基质细胞通过重编程造血干细胞Ng,J。J.; Marneth,A.E。;格里菲斯(Griffith) younge
是从MSC和CPG-ODN前基于MSC的调节培养基中开发的,其细菌清除率明显更高,而肺部感染后中性粒细胞粒细胞肉芽肿则比对照小鼠相比。在目标下进行切割,并使用核酸酶(切割和运行)染色质测序释放,我们识别出MSC条件的培养基在骨髓中涉及的基因和MTOR Pathway信号持久性的HSC中在HSC中留下H3K4ME3组蛋白标记。MSC的可溶性因子和细胞外囊泡介导了HSC上的这些OFECT和质量分析的蛋白质组学分析,这揭示了可溶性钙网蛋白作为潜在的培训。总而言之,这项研究表明,训练有素的免疫力可以由MSC的旁分泌因子介导,从而通过对中性粒细胞介导的抗菌抗菌免疫的长期功能变化来诱导嗜中性粒细胞训练的免疫力。
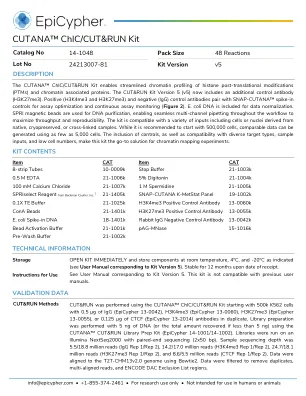
14-1048 技术数据表
CUTANA™ChIC/CUT&RUN 试剂盒可简化组蛋白翻译后修饰 (PTM) 和染色质相关蛋白的染色质分析。CUT&RUN 试剂盒版本 5 (v5) 现在包含额外的对照抗体 (H3K27me3)。阳性 (H3K4me3 和 H3K27me3) 和阴性 (IgG) 对照抗体与 SNAP-CUTANA™spike-in 对照配对,用于优化测定和持续测定监测 (图 2)。包含大肠杆菌 DNA 以进行数据标准化。SPRI 磁珠用于 DNA 纯化,可在整个工作流程中实现无缝多通道移液,从而最大限度地提高通量和可重复性。该试剂盒与各种输入兼容,包括源自天然、冷冻保存或交联样本的细胞或细胞核。虽然建议从 500,000 个细胞开始,但只需 5,000 个细胞即可生成可比数据。纳入对照以及与多种目标类型、样本输入和低细胞数量的兼容性,使该试剂盒成为染色质映射实验的首选解决方案。

COMPASS 亚基 Spp1 通过限制 DNA 对核酸酶的利用来保护 Tus/Ter 复制叉屏障处的新生 DNA
同源重组因子在 DNA 复制过程中对保护新生 DNA 起着至关重要的作用,但染色质在此过程中的作用尚不清楚。在这里,我们使用了已知可在酿酒酵母中诱导位点特异性复制叉停滞的细菌 Tus/Ter 屏障。我们报告称,Set1C 亚基 Spp1 被募集到停滞的复制叉后面,与其与 Set1 的相互作用无关。Spp1 染色质募集依赖于其 PHD 结构域与沉积在停滞叉后面的 H3K4me3 亲本组蛋白的相互作用。它的募集通过限制 Exo1 的访问来防止 ssDNA 在停滞叉处积累。我们进一步表明,删除 SPP 1 会增加屏障上游的突变率,有利于微缺失的积累。最后,我们报告称 Spp1 保护 Tus/Ter 停滞复制叉处的新生 DNA。我们认为 Spp1 限制了叉的重塑,最终限制了新生 DNA 对核酸酶的利用。

急性髓样白血病中的Smad1损失KMT2A
结果:在这里,我们报告了KMT2A :: AFF1和KMT2A :: MLLT3融合基因依赖性基因的下调Smad1(TGF-B信号轴转录因子)。SMAD1表达在大多数AML患者样品和包含两个融合基因KMT2A :: AFF1和KMT2A :: MLLT3的细胞系中丢失。SMAD1表达的丧失是通过将两个KMT2A融合基因引入造血干细胞和祖细胞中的。SMAD1的损失与具有KMT2A :: AFF1和KMT2A :: MLLT3的测试细胞中SMAD1启动子的H3K4me3水平显着降低。Smad1在具有KMT2A :: AFF1融合基因的细胞中的表达影响了细胞在体外的体外和影响kmt2a :: aff1细胞系MV4-11的体外植入。在MV4-11细胞中Smad1表达引起Hoxa9和Meis1的下调,这是通过TGF-B刺激加强的。 此外,在MV4-11细胞中,SMAD1的存在敏化细胞对TGF-B介导的G1暂停。在MV4-11细胞中Smad1表达引起Hoxa9和Meis1的下调,这是通过TGF-B刺激加强的。此外,在MV4-11细胞中,SMAD1的存在敏化细胞对TGF-B介导的G1暂停。

启动子的全基因组核小体分辨率图
摘要 增强子-启动子环路模型长期以来一直主导着基因调控领域,其中增强子通过物理接触激活其靶基因。然而,由于存在替代机制的证据以及缺乏系统验证(主要是由于缺乏合适的实验技术),该模型的普遍性受到了质疑。在本研究中,我们提出了一种新的基于 MNase 的邻近连接方法,称为 MChIP-C,该方法可以在基因组范围内以单核小体分辨率测量蛋白质介导的染色质相互作用。通过应用 MChIP-C 研究 K562 细胞中以 H3K4me3 启动子为中心的相互作用,我们发现与基于限制性内切酶的 C 方法相比,它具有大大提高的分辨率和灵敏度。这使我们能够将 EP300 组蛋白乙酰转移酶和 SWI/SNF 重塑复合物确定为建立和/或维持增强子-启动子相互作用的潜在候选者。最后,利用已发表的 CRISPRi 筛选数据,我们发现大多数经过功能验证的增强子确实与其同源启动子发生物理相互作用,支持增强子-启动子环路模型。

跨学科研究:研究人员职业跨学科研究的动机和挑战
转座元素(TES)是寄生虫DNA序列,能够沿所有基因组的染色体移动和繁殖。可以通过靶向沉默表观遗传标记来控制它们,这可能会影响包括基因在内的相邻序列的Chro Matin结构。在这项研究中,我们使用了来自几个果蝇Melanogaster的卵巢样品和果蝇Simulans野生型菌株产生的转录组和表观基因组高吞吐量数据,以精细量化Te插入对基因RNA水平和组蛋白标记的影响(H3K9ME3和H3K9ME3和H3K4ME3)。我们的结果揭示了与梅拉·诺加斯特(D. Mela Nogaster)相比,TES对D. simulans中直源基因的表观遗传作用更强。同时,我们发现了D. mel Anogaster基因组中TE对基因H3K9me3的差异的较大贡献,这证明了Te数字周围的Te数与D. melanogaster中这种染色质标记的水平的更强相关性。总体而言,这项工作有助于理解TE在基因组中的物种特异性影响。它为TE提供的可观自然变异性提供了新的启示,这可能与适应性和进化潜力的对比有关。

糖尿病和高脂血症中的免疫训练
缩写:乙酰辅酶 A,乙酰辅酶 A;ASCVD,动脉粥样硬化性心血管疾病;ATM,脂肪组织巨噬细胞;BCG,卡介苗;CRP,高敏 C 反应蛋白;DAMP,损伤相关分子模式;FH,富马酸水合酶;H3K27ac,组蛋白 3 赖氨酸 27 乙酰化;H3K4me1,组蛋白 3 赖氨酸 4 单甲基化;H3K4me3,组蛋白 3 赖氨酸 4 三甲基化;HIF1 α,缺氧诱导因子 1 α;HITI,高血糖诱导的训练免疫;IL-1 β,白细胞介素 1 β;IL-6,白细胞介素 6;Ldlr,低密度脂蛋白受体; Lp(a),脂蛋白(a);LPS,脂多糖;LXRs,肝脏X受体;mTOR,雷帕霉素的机制靶点;NK,自然杀伤细胞;oxLDL,氧化LDL;OxPLs,氧化磷脂;PAMPs,病原体相关分子模式;PBMCs,外周血单核细胞;PRRs,模式识别受体;SAT,皮下脂肪组织;TCA,三羧酸循环;TIH,短暂性间歇性高血糖症;TLR,Toll样受体;TNF-α,肿瘤坏死因子α;VAT,内脏脂肪组织;WD,西方饮食。