XiaoMi-AI文件搜索系统
World File Search SystemHRAS

AR+、HRAS/PIK3CA 共突变涎管癌病例系列的靶向治疗
结果:从 MTB 中确定了 4 名具有 AR+ HRAS / PIK3CA 共突变 SDC 和临床随访数据的患者。从文献中确定了另外 9 名具有临床随访的患者。除了 AR 过表达和 HRAS 和 PIK3CA 变异之外,还确定了 PD-L1 表达和肿瘤突变负荷 > 每兆碱基 10 个突变作为其他潜在可靶向变异。在可评估的患者中,7 名患者开始接受雄激素剥夺疗法 (ADT)(1 名部分缓解 (PR)、2 名稳定疾病 (SD)、3 名进展性疾病 (PD)、2 名不可评估),6 名患者开始接受替比法尼治疗(1 名 PR、4 名 SD、1 名 PD)。各有一名患者接受了免疫检查点抑制(混合反应)以及替比法尼和 ADT(SD)以及阿哌沙布和 ADT(PR)的联合疗法治疗。

Tipifarnib 治疗复发性、转移性 HRAS 突变型涎腺癌
通讯作者:Alan L. Ho,医学博士,哲学博士,纽约纪念斯隆凯特琳癌症中心血液学/肿瘤学系,1275 York Ave,纽约,NY 10065(hoa@mskcc.org)。作者贡献 Glenn J. Hanna:研究概念和设计、数据采集、数据分析和解释、撰写初稿以及撰写审阅和编辑。Jeffrey P. Guenette:数据管理、图像分析以及撰写审阅和编辑。Nicole G. Chau、Cyrus M. Sayehli、Christian Wilhelm、Robert Metcalf、Deborah J. Wong、Marcia Brose、Mohammad Razaq、Elisabeth Pérez-Ruiz、Ezra EW Cohen 和 Rahul Aggarwal:数据采集、数据解释以及撰写审阅和编辑。 Catherine Scholz 和 Antonio Gualberto:方法论、药物赞助、项目管理以及写作-审查和编辑。Alan L. Ho:概念化、设计、数据采集、数据分析和解释以及写作-审查和编辑。
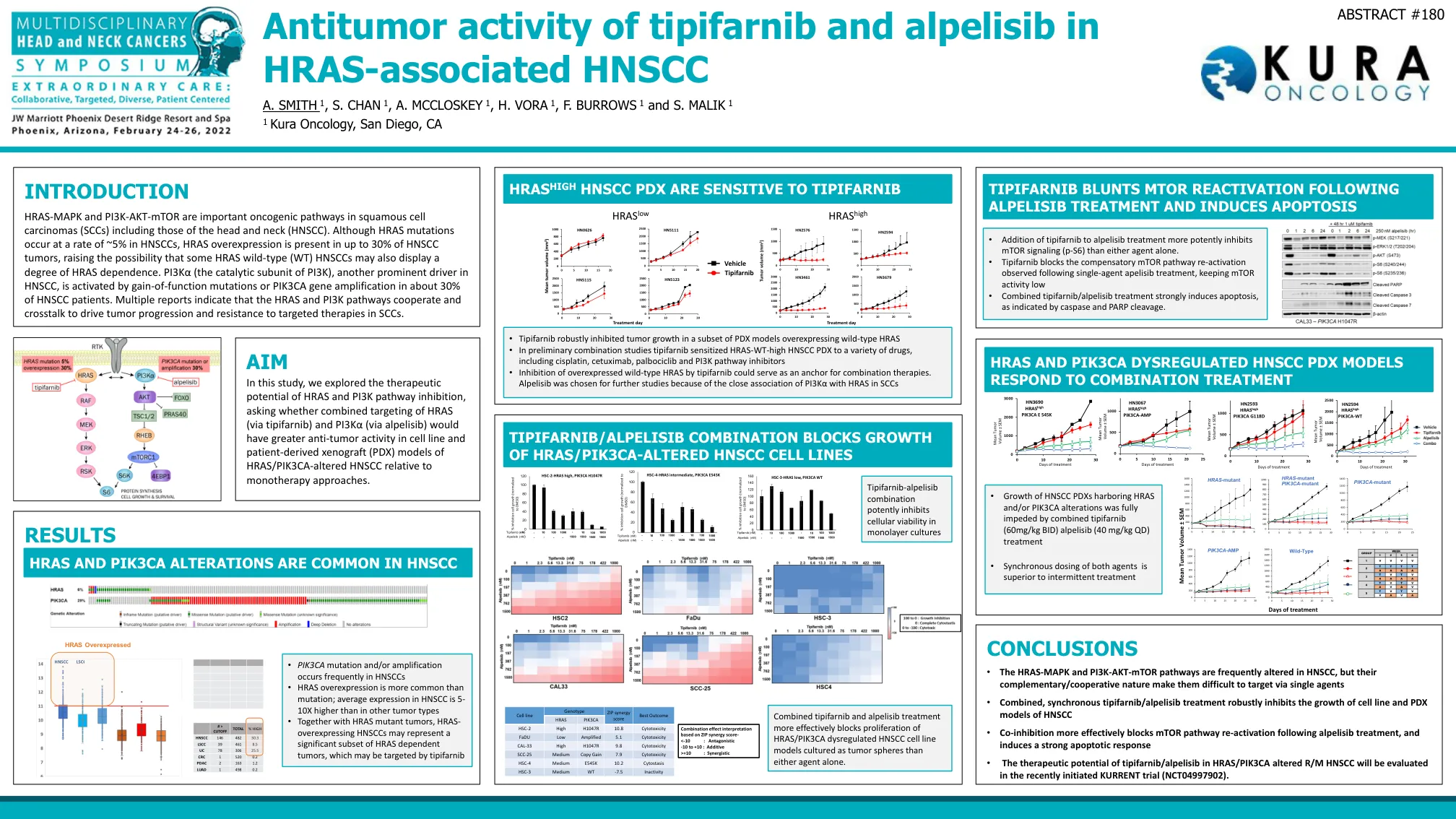
引言 结论 目的 结果
HRAS-MAPK 和 PI3K-AKT-mTOR 是鳞状细胞癌 (SCC)(包括头颈部鳞状细胞癌 (HNSCC))中的重要致癌通路。尽管 HRAS 突变在 HNSCC 中的发生率约为 5%,但 HRAS 过表达在多达 30% 的 HNSCC 肿瘤中存在,这增加了一些 HRAS 野生型 (WT) HNSCC 也可能表现出一定程度的 HRAS 依赖性的可能性。PI3Kα(PI3K 的催化亚基)是 HNSCC 中的另一个主要驱动因素,在约 30% 的 HNSCC 患者中,PI3Kα 被功能获得性突变或 PIK3CA 基因扩增激活。多份报告表明,HRAS 和 PI3K 通路协同和串扰,以驱动 SCC 中的肿瘤进展和对靶向疗法的耐药性。

订单 – 分子病理学/免疫组织化学
NGS小组,FFPE融合分析(RNA),FFPE☐结肠(BRAF/KRAS/NRAS/PIK3CA)☐肺(ALK,ROS1和RET)☐CLL突变分析☐肺(BRAF/kras/kras/nras/nras/pik3ca/fuson and eff in nraver/exnaly nrative) IST(BRAF/KIT/PDGFRA- POSIGNA/PAM50☐igh☐黑色素瘤(BRAF/KIT/NRAS/HRAS)(子类别和风险得分)☐TCR☐MTC(BRAF/KRAS/HRAS/HRAS/RET) BRCA2/PIK3CA)☐LOH1P19Q (包括 IDH1/2- ☐ B-ALL ☐ 乳腺(PIK3CA)突变分析),诊断时的 FFPE 母细胞比例:NGS 面板,cfDNA/血浆 ☐ TERT 启动子突变分析 ☐ MRD 随访,☐ 肺(EGFR/KRAS/NRAS/BRAF)(c.1-124C>T / c.1-146C>T)天:

rmc-6236,一个RAS(ON)多选择性三重复合抑制剂
(1)基于基础医学洞察力的QUSON,NRA和HRAS突变的频率G12X,G13X和Q61X在2022年3月3日的实体瘤中估计的百分比
量化英语学校的过热风险 - UCL Discovery
PMVK 4,NSDHL,4 HRAS 4和KRT10。4到目前为止的疾病机制包括种系X连锁变体,镶嵌变体和种系首次击中,并以马赛克第二击中命中。配对的血液和受影响的皮肤DNA接受了深层外显子组测序(WES,平均250倍),N¼14,如果阴性为阴性,皮肤DNA经过了靶向测序板R327(Mosaic Disorsisters R327)(UK National Genomic Test Directory),N¼8。两名患者因样本限制而没有前进到下一代测序面板。在研究后期招募的两名患者首先是下一代测序小组,但没有继续进行WES。我们在这里确定ilven具有多种单基因原因,在NSDHL中发生突变(N¼2,种系,NSDHL C.613G [t,p。[G205T],C.603_604DELTG,p。[H201FS*69],no no smine,pmvk(no smine),pmvk(no),在皮肤中的同一基因中检测到的变体PMVK C.126delg,P.R42Fs,在WES上拾取),HRAS(N¼1,Mosaic,Hras C.37G [C,p。(G13R),在面板上拾取,和Card14(N¼2,Mosaic,this 2,cocaic of this 2 coped of trapered of。十名患者没有鉴定出病原变体,我们特别排除了所有先前描述的基因中的任何变体。没有对WES阴性的患者在随后的面板上鉴定出的基因,这表明尚未确定的变体不是已知的镶嵌基因,或者如果它们不太可能是

rmc-6236,一个RAS(ON)多选择性三重复合抑制剂
(1)基于基础医学洞察力的QUSON,NRA和HRAS突变的频率G12X,G13X和Q61X在2022年3月3日的实体瘤中估计的百分比

第 18 章 RAS 致癌基因的故事
第 18 章 RAS 致癌基因的故事 221007bu3 抗癌药物:发现和寻求治愈方法的故事 Kurt W. Kohn,医学博士,哲学博士 名誉科学家 分子药理学实验室 发育治疗学分部 美国国立癌症研究所 马里兰州贝塞斯达 kohnk@nih.gov 第 18 章 RAS 致癌基因的故事 病毒中的 RAS 致癌基因。RAS 基因是人类癌症中一个特别重要的基因或致癌基因家族,它首次是在对致癌病毒的研究中发现的。1963 年的某个时候,在伦敦医院研究实验室癌症研究部工作的 Jennifer Harvey 给小鼠和大鼠接种了一只患有病毒诱发的白血病的大鼠的血浆。她定期将病毒从一只动物转移到另一只动物,从而诱发它们患上白血病。然而,那一年的一次,她注意到一些不寻常的东西,这为癌症的成因和治疗打开了一扇新的窗户(Harvey,1964 年)。接种了她一只白血病大鼠病毒的小鼠,除了常见的白血病(血液和淋巴结中有恶性细胞,而不是各种组织中的肿块)外,还意外地患上了实体瘤。后来发现,她的白血病病毒从大鼠自己的基因组中获取了一段 DNA 片段(拼接到其基因组中)。这段 DNA 现在是新病毒基因组的一部分,导致她的小鼠出现实体瘤型癌症肿块。此外,新的癌症基因被发现是正常基因 RAS 的突变版本(可能是大鼠肉瘤,突变版本最早是在大鼠肉瘤中发现的)。Harvey 的名字因新发现的 HRAS 致癌基因中的字母 H 而永垂不朽,HRAS 致癌基因是正常 HRAS 基因的突变形式。哈维的新病毒导致培养皿表面的细胞过度生长,形成“病灶”(图 18.1),其方式与温伯格团队后来在致癌基因研究中观察到的情况类似(第 15 章中的图 15.3)。电子显微镜图像中看到的哈维病毒颗粒具有非常不寻常的结构,类似于辐条轮(图 18.2)。

卫生福利部食品药物管理署精准医疗分子检测实验室列册...
ABL1 AKT1 AKT3 ALK AR AXL BRAF CCND1 CDK4 CDK6 CTNNB1 DDR2 EGFR ERBB2 ERBB2 ERBB3 ERBB3 ERG ESR1 ESR1 ETV1 ETV1 ETV4 ETV4 ETV5 ETV5 ETV5 ETV5 FGFR1 MAP2K2 MET MTOR MYC MYCN NRAS NTRK1 NTRK2 NTRK3 PDGFRA PIK3CA PPARG RAF1 RAS ROS1 SMO

人为可靠性分析 (HRA) 简介
THERP 修订,ASEP 生成;新的模拟模型;对核电行业安全性和可靠性的关注(例如 TMI);标准化的 HRA 流程;新的 HRA 数据库;新的专家评估技术;HRA 在风险评估中的整合度不断提高。切尔诺贝利事故体现了人为失误在灾难中的作用。恢复工作已解决。建模框架 — Rasmussen:S、R 和 K;原因:失误、疏忽和错误;时间可靠性相关性;引入绩效塑造因素 (PSF)