XiaoMi-AI文件搜索系统
World File Search SystemHoffmann
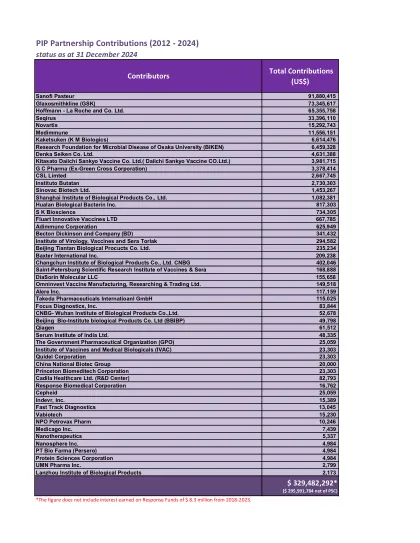
PIP 合作伙伴贡献(2012 - 2024)
赛诺菲巴斯德 91,880,415 葛兰素史克(GSK) 73,345,617 霍夫曼-罗氏公司 65,355,758 Seqirus 33,396,110 诺华 15,292,743 Medimmune 11,556,151 Kaketsuken(KM Biologics) 6,614,476 大阪大学微生物疾病研究基金会(BIKEN) 6,459,328 Denka Seiken Co. Ltd. 4,631,388 北里第一三共疫苗有限公司(Daiichi Sankyo Vaccine CO.Ltd.) 3,981,715 GC Pharma(前绿十字公司) 3,378,414 CSL Limted 2,667,745布塔坦研究所 2,730,303 科兴生物制品有限公司 1,453,267 上海生物制品研究所有限公司 1,082,381 华兰生物疫苗股份有限公司 817,303 SK Bioscience 734,305 Fluart Innovative Vaccines LTD 667,785 Adimmune Corporation 625,949 Becton Dickinson and Company (BD) 341,432 病毒、疫苗和血清研究所 Torlak 294,582 北京天坛生物制品股份有限公司 235,234 百特国际公司 209,238 长春生物制品研究所有限公司 CNBG 402,046 圣彼得堡疫苗和血清科学研究所 168,888 DiaSorin Molecular LLC 155,658 Omninvest Vaccine Manufacturing, Researching & Trading Ltd. 149,518 Alere Inc. 117,159 Takeda Pharmaceuticals Internatioanl GmbH 115,025 Focus Diagnostics, Inc. 83,844 CNBG-武汉生物制品研究所有限公司52,678 北京生物研究所生物制品有限公司(BBIBP) 49,798 Qiagen 61,512 印度血清研究所有限公司 48,335 政府制药组织(GPO) 25,059 印度疫苗和医学生物制品研究所(IVAC) 23,303 Quidel Corporation 23,303 中国生物技术集团 20,000 Princeton Biomeditech Corporation 23,303 卡迪拉医疗保健有限公司(研发中心) 82,793 Response Biomedical Corporation 16,762 Cepheid 25,059 Indevr, Inc. 15,389 Fast Track Diagnostics 13,045 Vabiotech 15,230 NPO Petrovax Pharm 10,246 Medicago Inc. 7,439 Nanotherapeutics 5,337 Nanosphere Inc. 4,984 PT Bio Farma (Persero) 4,984 Protein Sciences Corporation 4,984 UMN Pharma Inc. 2,799 兰州生物制品研究所 2,173

wenn dna警报auslöst
背景信息•背景信息•背景信息背景信息,用于分配Paul Ehrlich-和Ludwig Darmstaedter奖2025年教授博士。 Andrea Ablasser,博士教授格伦·巴伯(Glen Barber)和博士教授当DNA警报触发我们身体的细胞时,Zhijian J. Chen暴露于许多不同的威胁。这包括例如病毒感染,癌症和其发电厂(线粒体)中的事故。所有这些威胁共同表明它们在没有生意的细胞等离子体中显示了DNA双链(DSDNA)。那里信号外国遗传信息。也不应出现在细胞核和线粒体之外。随着我们先天的免疫系统承认并消除了错误位置的DNA的危险,长期以来一直是一个谜。这三名获奖者在2008年至2013年之间解决了这一问题,从那以后,它得到了越来越广泛的通知。他们在开头发现了一个信号路径,酶传感器为。一旦他在细胞等离子体中跟踪dsdna,他就会抓住她。这会改变其形状,从而可以催化分子信使的形成。该使者控制着一个细胞内受体,该受体通过使某些基因对齐在细胞核中接受并转换信使的信息:立即产生干扰素。这些干扰素散布在周围的组织中,并寻求帮助。这违反了我们的免疫系统“奇怪”和“本身”必须明确区分的规则。区别于所谓的CGAS-sting-Pathway是其普遍性:它的传感器没有区分外部和人体自己的DSDNA。这种违反规则的行为是有风险的,因为它具有无意自我毁灭的可能性。它提供了一种双重方法来干预此信号路径。每天我们受到数千种细菌和病毒的攻击。在大多数情况下,我们的身体成功地抵御了这些攻击。这要归功于其先天的免疫系统,入侵者在国际象棋中持有它,直到他的信号激活了获得的免疫系统,抗体和T细胞以关闭攻击者。在此之前可能需要几天。没有天生的免疫力,如今我们几乎无法生存。尽管如此,他们的研究长期以来一直在阴暗的存在。虽然20th世纪非常精确地知道,很长一段时间以来,先天性免疫系统如何感知微生物攻击。仅通过朱尔斯·霍夫曼(Jules Hoffmann)和布鲁斯·贝特勒(Bruce Beutler)的发现而改变

德国研究、创新和技术绩效报告
阿部洋子(Yoko Abe)教授A.S. 博士鲍里斯·奥古斯基博士斯蒂芬妮·鲍尔博士Ingo Baumann、Marc Becker、教授A.S. 博士霍尔格·博宁 (Holger Bonin)、阿方斯·博托夫 (Alphonse Botthof) 博士Tanja Bratan,博士h. c. Edelgard Bulmahn,教授Theresa Cho 博士D.,教授Moon Choi 博士D.,博士安娜·克里斯特曼博士Jano Costard、Susanne Dehmel、Peter Dortans、Judith Ebel、John Elling、Gerhard Fasol 博士。 D.,教授A.S. 博士安妮特·弗兰克,教授A.S. 博士Volker Gass、Ronald Grasmann、Christoph Gross、Minoru Hanakat、Magnus Harviden、Prof. A.S. 博士Justus Haucap 博士克里斯蒂安·海德克 (Christian Heideck)、萨沙·赫尔曼 (Sascha Hermann);亚历山大·赫希菲尔德(Alexander Hirschfeld)教授A.S. 博士凯瑟琳娜·霍茨勒 (Katharina Hölzle) 博士托拜厄斯·霍夫曼博士Stefan Rolf Huebner,教授A.S. 博士Christian Hummert,教授饭岛克也博士D.,教授井地智宏博士D.,教授梶川雄也博士D.,博士Matthias Kautt、Jonas Kellner 博士金东益教授Hannah Kim 博士D.,教授So Young Kim 博士D.,杰西卡·金; Osamu Kobayashi 博士、Martin Kölling、Azusa Kondo 博士John König、Harald Conrad、Max Kroymann、Eddy Kwon 博士D.,博士Taek-ryoun Kwon 博士D.,Jae Hong Lee,博士D.,Joohyung Lee,博士D.,博士Sunghee Lee、Yonsoo Lee、Youngmin Lee 博士D.,博士Jochen Legewie 博士、Eckart Lilienthal、Andreas Lindenthal 博士Johannes Ludewig 博士,Valeska Maul;洛塔尔·门尼肯博士乔治·梅茨格博士Susanne Meyer,教授A.S. 博士Paul P. Momtaz,教授A.S. 博士克劳迪娅·穆勒 (Claudia Muller) 教授A.S. 博士长野宏司教授长冈贞夫博士D.,教授Alice Oh,博士Kazuaki Osumi 博士,PD Anne Otto,Byeongwon Park,博士D.,博士Byeungkwan Park 教授、Jung Ho Park 教授Sangook Park 博士D. 太阳

GaN 高电子迁移率晶体管外延堆栈中穿线位错引起的垂直场不均匀性
数据可用性声明:支持本研究结果的数据可根据合理要求从通讯作者处获取。1 H. Amano、Y. Baines、E. Beam 等人,2018 年 GaN 电力电子路线图,Journal of Physics D: Applied Physics。51,(2018)。2 K. Husna Hamza 和 D. Nirmal,GaN HEMT 宽带功率放大器综述,AEU - 国际电子和通信杂志。116,153040 (2020)。3 G. Meneghesso、M. Meneghini、I. Rossetto、D. Bisi、S. Stoffels、M. Van Hove、S. Decoutere 和 E. Zanoni,GaN 基功率 HEMT 的可靠性和寄生问题:综述,半导体科学与技术。31,(2016)。 4 JA del Alamo 和 J. Joh,GaN HEMT 可靠性,微电子可靠性。49,1200-1206 页 (2009)。5 M. Meneghini、A. Tajalli、P. Moens、A. Banerjee、E. Zanoni 和 G. Meneghesso,基于 GaN 的功率 HEMT 中的捕获现象和退化机制,半导体加工材料科学。78,118-126 页 (2018)。6 B. Kim、D. Moon、K. Joo、S. Oh、YK Lee、Y. Park、Y. Nanishi 和 E. Yoon,通过导电原子力显微镜研究 n-GaN 中的漏电流路径,应用物理快报。104,(2014)。 7 M. Knetzger、E. Meissner、J. Derluyn、M. Germain 和 J. Friedrich,《用于电力电子的碳掺杂变化与硅基氮化镓垂直击穿之间的关系》,《微电子可靠性》。66,16-21 (2016)。 8 A. Lesnik、MP Hoffmann、A. Fariza、J. Bläsing、H. Witte、P. Veit、F. Hörich、C. Berger、J. Hennig、A. Dadgar 和 A. Strittmatter,《碳掺杂氮化镓的性质,固体物理状态 (b)》。254,(2017)。 9 B. Heying、EJ Tarsa、CR Elsass、P. Fini、SP DenBaars 和 JS Speck,《位错介导的氮化镓表面形貌》,《应用物理学杂志》。 85,6470-6476 (1999)。

向风湿病学家学习什么?
1 Cron, RQ 和 Chatham, WW《风湿病学家在 Covid-19 中的作用》。《风湿病学杂志》。2020 年。47 (5) 639-642。2 Misra DP、Agarwal V、Gasparyan AY、Zimba O。风湿病学家对冠状病毒病 19 (COVID-19) 和潜在治疗靶点的看法 [2020 年 4 月 10 日提前在线发表]。《临床风湿病学》。2020;1 - 8。3 Hoffmann M、Kleine-Weber H、Schroeder S 等人。SARS-CoV-2 细胞进入依赖于 ACE2 和 TMPRSS2,并被临床验证的蛋白酶抑制剂阻断。《细胞》。2020;181(2):271 - 280.e8。 4 Mahevas, M. 等人 (2020)。没有证据表明羟氯喹对需要氧气的 COVID-19 感染住院患者有临床疗效:使用常规收集的数据模拟目标试验的研究结果。medRxiv。5 Huang C、Wang Y、Li X 等人。中国武汉 2019 年新型冠状病毒感染患者的临床特征。柳叶刀。2020;395(10223):497 ‐ 506。6 Williamson, B.、Feldmann, F.、Schwarz, B.、Meade-White, K.、Porter, D.、Schulz, J.、...... 和 Okumura, A. (2020)。瑞德西韦对感染 SARS-CoV-2 的恒河猴的临床益处。BioRxiv。 7 Chen, Z., Hu, J., Zhang, Z., Jiang, S., Han, S., Yan, D., ... & Zhang, Z. (2020). 羟氯喹对 COVID-19 患者的疗效:一项随机临床试验的结果。MedRxiv。8 Cantini F、Niccoli L、Matarrese D、Nicastri E、Stobbione P、Goletti D。巴瑞替尼治疗 COVID-19:安全性和临床影响的初步研究。J Infect。2020;S0163-4453(20)30228-0。9 Xu, X.、Han, M.、Li, T.、Sun, W.、Wang, D.、Fu, B., ... & Zhang, X. (2020). 托珠单抗对重症 COVID-19 患者的有效治疗。ChinaXiv,202003 (00026),V1。 10 Monti S, Balduzzi S, Delvino P, Bellis E, Quadrelli VS, Montecucco C. 接受免疫抑制靶向疗法治疗的一系列慢性关节炎患者的 COVID-19 临床病程。Ann Rheum Dis. 2020;79(5):667 ‐ 668。

针对 SARS-CoV-2 的进入步骤
当前,由 SARS-CoV-2 引起的 COVID-19 正在人类中迅速传播,构成全球卫生紧急情况(https://www.who.int/);截至 2020 年 4 月 29 日,全球确诊病例 3,018,681 例,死亡 207,973 例。了解冠状病毒的受体识别机制,从而调整其致病机制、传播速度和宿主范围,是战胜这一流行病的关键。1,2 冠状病毒的 S 蛋白是病毒入侵细胞所必需的。此外,进入需要细胞蛋白酶来引发 S 蛋白;它们在 S1/S2 和 S2' 位点裂解 S 蛋白,从而促进由 S2 亚基介导的病毒和靶细胞膜的融合。众所周知,SARS-CoV以血管紧张素转换酶2(ACE2)作为进入受体,并利用细胞丝氨酸蛋白酶TMPRSS2来启动S蛋白。3,4 SARS-2-S与SARS-S之间的氨基酸同源性约为76%,1但SARS-CoV-2如何进入仍有待充分探索。SARS-2-S与SARS之间的氨基酸同源性为了进一步了解病毒进入的机制,Hoffmann等人首先寻找SARS-2-S有效蛋白水解的证据。带有C端抗原标签的293T细胞表达的SARS-2-S蛋白的免疫印迹分析显示一条S2亚基带,表明SARS-2-S可以被有效水解,这与其S1 / S2裂解位点存在几个精氨酸残基相一致。有趣的是,冠状病毒的人畜共患潜力是由 S 蛋白的裂解位点序列决定的。1 因此,还需要进一步研究以了解 SARS-CoV-2 入侵人细胞是否也需要多碱基裂解位点,并详细描述这些裂解位点。接下来,作者使用携带 SARS-2-S 和 SARS-S 的 VSV 病毒感染一系列人和动物细胞系,并观察到它们侵入相同的细胞谱系。与这一发现一致的是,ACE2 和 SARS-S 结合所必需的大多数氨基酸在 SARS-2-S 中是保守的,并且定向表达 ACE2,而不是人 DPP4 或人氨基肽酶 N(MERS-CoV 和 HCoV-229E 的进入受体),使得 SARS-CoV-2 和 SARS-CoV 能够成功感染不敏感的 BHK-21 细胞。此外,针对人 ACE2 产生的抗血清可以保护 BHK-21 细胞免受 SARS-CoV-2 和 SARS-CoV 的侵袭。总而言之,这些研究强烈暗示 ACE2 是 SARS-CoV-2 的细胞受体。在

数据表:MCA1396D550产品详细信息
参考文献1。Els Conrath,K。等。(2001)骆驼单域抗体作为双特异性和双价抗体构建体中的模块化建筑单元。J Biol Chem。 276(10):7346-50。 2。 suen,J.L。 等。 (2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。 免疫。 103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。276(10):7346-50。2。suen,J.L。等。(2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。免疫。103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。103:301-309。3。Hoffmann,S.C。等。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。282(31):22370-5。4。Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Zheng,J。等。(2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。BMC免疫。8:4。5。Bahi,A。&Dreyer,J.L。(2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。基因脑行为。7(2):244-56。6。Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Wrighton,K.H。等。(2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。284(15):9755-63。7。Diefenbacher,M。等。(2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。8。J Biol Chem。 286:25027-38。 Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放J Biol Chem。286:25027-38。Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Alvarez,M.M。等。(2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放PLOS ONE。5:E10176。9。Bahi,A。等。(2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。神经心理药理学。33:2726-34。10。Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Gunnarsen,K.S。等。(2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放BMC生物技术。10:8。11。Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Hwang,H.Y。等。(2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。mol免疫。45:2570-80。12。de Vooght,L。等。(2012)功能性的表达和细胞外释放

大疱性表皮松解症诊断的基因组
关于 EB 的诊断和分类。 J Am Acad Dermatol. 2008;58:931---50。 6. Oliveira ZN、Périgo AM、Fukumori LM 和Aoki V. 遗传性大疱性表皮松解症的免疫学映射。胸罩皮肤科。 2010;85:856---61。 7. Has C, He Y.研究技术变得简单:大疱性表皮松解症的免疫荧光抗原图谱。 J Invest Dermatol。 2016;136:e65---71。 8. Takeichi T、Liu L、Fong K、Ozoemena L、McMillan JR、Salam A 等人。全外显子组测序提高了诊断性大疱性表皮松解症实验室的突变检测能力。 Br J 皮肤病学。 2015;172:94---100。 9. Tenedini E、Artuso L、Bernardis I、Artusi V、Percesepe A、De Rosa L 等。基于扩增子的下一代测序:大疱性表皮松解症分子诊断的有效方法。 Br J 皮肤病学。 2015;173:731---8。 10. Has C、Küsel J、Reimer A、Hoffmann J、Schauer F、Zimmer A 等。靶向二代测序在大疱性表皮松解症诊断中的地位。 Acta Derm Venereol。 2018;98:437---40。 11. Vahidnezhad H、Youssefian L、Saeidian AH、Touati A、Sotoudeh S、Abiri M 等人。多基因下一代测序面板可识别患有未知亚型大疱性表皮松解症的患者的致病变异:具有预后意义的亚分类。 J Invest Dermatol。 2017;137:2649---52。 12. Lucky AW、Dagaonkar N、Lammers K、Husami A、Kissell D 和 Zhang K. 一种用于诊断大疱性表皮松解症的综合下一代测序检测方法。小儿皮肤病学。 2018;35:188---97。 13. Mariath LM、Santin JT、Frantz JA、Doriqui MJR、Kiszewski AE、Schuler-Faccini L. 巴西大疱性表皮松解症的遗传基础概述:发现新的和复发的致病变异。临床遗传学。 2019;96:189---98。 14.Yiasemides E、Walton J、Marr P、Villanueva EV、Murrell DF。透射电子显微镜与免疫荧光成像在大疱性表皮松解症诊断中的对比研究。 Am J Dermatopathol。 2006;28:387---94。 15. Saunderson RB、Vekic DA、Mallitt K、Mahon C、Robertson SJ、Wargon O. 一项回顾性队列研究,评估与免疫荧光和

90 分钟或更短时间内看世界:火箭物流和未来……
这篇研究文章在 2022 年 CJCS 国防和军事战略论文竞赛的战略研究论文类别中获得第二名。虽然 JFSC 的许多学生都取得了很高的研究水平,但提交给比赛的论文代表了专业军事教育机构每年完成的一些最好的研究、写作和思考。获得认可的手稿达到了极高的标准。其他比赛获奖者将在《联合部队季刊》上找到。由 1 CDR Von P. H. Fernandes、Maj Ashley Gunn、MAJ Lucas Hoffmann 和 Lt Col Nita McQuitery 撰写 1959 年,海军少将亨利·埃克尔斯 (Henry Eccles) 将后勤描述为“国家经济与其作战部队战术行动之间的桥梁”。 1 火箭后勤,即使用轨道级火箭将货物从地球上的一个地方运送到另一个地方,有可能大大缩短这一距离。自埃克尔斯发表声明以来的 70 年里,对更快、更高效的后勤行动的竞争需求改变了国防部 (DoD) 开展后勤的方式以及后勤对全球军事行动的贡献。高效、精简的后勤不仅为指挥官提供了行动自由,而且扩大了作战范围。国防部开展高效后勤行动的能力决定了美国在全球范围内投射力量的程度和范围。虽然空运成为第二次世界大战后勤行动的一个特色,但货船继续为全球军事行动运输绝大多数物资和军事装备。空运加快了后勤工作,但需要付出财政成本,并且对重量和体积有限制。然而,空运速度对于美国欧洲司令部、美国非洲司令部、美国南方司令部和美国印太司令部战区的一些后勤挑战来说仍然不够快。美国太空军有一个新的尖端研发项目,即先锋计划,该计划正在探索使用轨道级火箭进行点对点运输。火箭物流承诺比战场上的空运速度更快,但成本更高,对货物类型的限制也更多,在本文的范围内,货物类型定义为人员和设备。与民用运输任务不同,军事任务没有可预测的目的地,无法投资基础设施开发以确保顺利着陆。2因此,先锋计划寻求的解决方案是拥有全地形最终下降系统,具有坚固的外部结构以处理着陆时的异物碎片 (FOD),并使用新技术,例如目前由 NASA 开创的技术,这将允许下降的火箭在最终下降过程中创建着陆台。

当DNA触发警报
背景信息 - 背景信息 - 背景信息背景信息有关Paul Ehrlich和Ludwig Darmstaedter奖2025年授予的授予Andrea Ablasser教授,Glen Barber博士和Zhijian J. Chen教授的DNA时,DNA触发的牢房将使我们的身体的细胞暴露给许多不同的植物植物,包括病毒感染和癌症,癌症的牢房都暴露于我们的身体上,并将其触发。所有这些威胁的共同点是,它们会导致DNA双链(DsDNA)出现在细胞的血浆中 - 它们不属于它们,并且它们作为外国遗传信息的存在标志着最大的危险。即使我们自己的dsDNA也不应在细胞核和线粒体之外存在。我们先天的免疫系统如何承认和抵御错误位置的DNA危险,这是长期以来一直是一个谜。这三名获奖者在2008年至2013年之间解决了这一问题,此后越来越全面地澄清了它。他们发现了一种从酶传感器开始的信号通路,该传感器一旦在细胞质中检测到它,该传感器就会抓住DSDNA。酶传感器在过程中改变了其形状,从而使其能够催化分子信使的形成。此使者又触发了一个细胞内受体,该受体通过将自己的通信发送给细胞核中的某些基因,要求它们立即产生干扰素,从而接收和翻译使者的调度。这些干扰素扩散到周围的组织并寻求帮助。确实为医学提供了双重机会,可以在此信号通路中进行治疗。这种所谓的CGAS刺道途径的区别是它的普遍性:其传感器没有区分外源性DSDNA和内源性DSDNA。这违反了我们的免疫系统必须明确区分“外国”和“自我”的规则,这种违规行为更具风险,因为它具有无意的自我毁灭的可能性。我们每天都会受到数千种细菌和病毒的攻击。在大多数情况下,我们的身体成功地抵御了这些攻击。这要归功于其先天的免疫系统,它使入侵者保持远处,直到其信号激活了人体获得的免疫系统,后者的抗体和T细胞消除了攻击者,这可能需要几天的时间。没有天生的免疫力,如今我们几乎无法生存。尽管如此,对这种免疫力的研究长期以来一直带来了阴暗的存在。虽然在20世纪,详细阐明了获得的免疫力的基本特征,但长期以来一直尚不清楚先天免疫系统如何感知微生物攻击。这仅在1990年代中期发生了变化,这些发现是由朱尔斯·霍夫曼(Jules Hoffmann)独立进行的,这些发现没有获得免疫系统,而布鲁斯·贝特勒(Bruce Beutler)则在