XiaoMi-AI文件搜索系统
World File Search SystemHorvitz


秀丽隐杆线虫基因调控等位基因和记者抨击研究
基因调控是多细胞生物的重要过程,但识别功能性调控序列和机制可能具有挑战性。在秀丽隐杆线虫中,正向遗传学可以识别破坏生理过程的内源性突变(“等位基因”),从而以无偏见的方式定义功能序列(Brenner 1974;Trent、Wood 和 Horvitz 1988;Desai 等人 1988;Barton、Schedl 和 Kimble 1987)。基于 CRISPR 的基因组编辑可用于测试内源序列的功能和生理作用(Dickinson 和 Goldstein 2016;Vicencio 和 Cerón 2021)。报告基因检测中对非编码 DNA 进行系统性测试(例如“报告基因抨击”)可以识别功能序列,但不能直接检查生理功能(Aamodt、Chung 和 McGhee 1991;Didiano 和 Hobert 2006;Boulin、Etchberger 和 Hobert 2006;Nance 和 Frøkjær-Jensen 2019)。
micropub-biology-000241.pdf
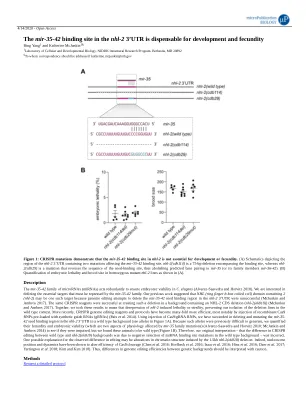
mir-35-42 家族的 microRNA (miRNA) 可重复发挥作用,以确保秀丽隐杆线虫的胚胎活力 (Alvarez-Saavedra 和 Horvitz 2010)。我们感兴趣的是确定 mir-35-42 家族必须抑制的必要靶标。我们之前的研究表明,NHL(环指 b-box 卷曲螺旋)结构域包含 2 (nhl-2) 可能就是这样一个靶标,因为基因组编辑尝试删除 nhl-2 3'UTR 中的 mir-35-42 种子结合区域失败 (McJunkin 和 Ambros 2017)。相同的 CRISPR 试剂在包含 NHL-2 CDS 缺失 (nhl-2(ok818)) 的背景下成功创建了这种缺失 (McJunkin 和 Ambros 2017)。总之,我们认为这些结果意味着 nhl-2 的去抑制会导致致死或不育,从而阻止我们在野生型背景下分离缺失系。最近,CRISPR 基因组编辑试剂和方案的效率提高了许多倍,最显著的是通过注射预装合成向导 RNA (gRNA) 的重组 Cas9 RNP(Paix 等人,2014 年)。通过注射 Cas9/gRNA RNP,我们成功地在野生型背景下删除和突变了 nhl-2 3'UTR 中的 mir-35-42 种子结合区(参见图 1A 中的等位基因)。由于此类等位基因以前难以生成,我们量化了它们的繁殖力和胚胎活力(这是受 mir-35 家族突变影响的两个生理方面)(Alvarez-Saavedra 和 Horvitz 2010;McJunkin 和 Ambros 2014),以查看它们是否受损,但我们发现这些动物是野生型(图 1B)。因此,我们最初的解释——野生型和 nhl-2(ok818) 背景之间的 CRISPR 编辑差异是由于野生型背景中 miRNA 结合位点突变的负选择——是不正确的。观察到的编辑差异的一个可能解释可能是 1.5kb nhl-2(ok818) 缺失引起的染色质结构改变。事实上,核小体的位置和动力学已被证明会改变 Cas9 切割的效率 (Chen 等人 2016;Horlbeck 等人 2016;Isaac 等人 2016;Hinz 等人 2016;Daer 等人 2017;Yarrington 等人 2018;Kim and Kim 2018)。因此,应谨慎解读不同遗传背景之间基因组编辑效率的差异。

PCAST 会议 - 白宫
Error 500 (Server Error)!!1500.That’s an error.There was an error. Please try again later.That’s all we know.

22-23 年度报告
萨曼莎·德拉赫 温迪、吉姆和埃塔·德雷珀 安·德罗斯特 安东尼娅·埃利斯 托尼亚·埃利斯 吉恩·爱泼斯坦 约瑟夫和桑德拉·法库尔特基金会 威廉和凯瑟琳·E.·费伦巴赫 Jr. 妮可·费拉罗 贝弗利·菲奇 钱恩·福勒-斯佩尔曼 伊莲·芬·弗罗恩吉安 黛博拉·弗鲁茨 罗伯特·富尔斯特和路易斯·艾布拉姆斯 帕特里克·甘农 杰曼·吉比安 凯瑟琳·吉格里奥蒂 杰西卡·吉格里奥蒂 凯莉·吉格里奥蒂 保罗·吉格里奥蒂 安和罗伯特·吉莱斯皮 麦肯纳·格洛里奥索 梅丽莎·格洛里奥索 埃尔伯特·古德 帕梅拉和艾迪森·古德尔慈善基金会 雪莉·戈登 邦妮·古尔利 詹姆斯·格雷厄姆 凯莉·格雷森 莎拉·格雷维特 艾比·格里菲斯 艾伦·哈尔丰 布里·汉考克 亨利·汉森 詹妮弗·哈里斯 贝琪·哈廷和 David R. Hexter 捐赠者建议基金 Doug 和 Suzanne Hicks John 和 Lisa Hiland John Hildebrand Cynthia A Hill 女士 Julius Hill Julia Horvath Michael J. Horvitz 2 捐赠者建议基金 Gabrielle Howard

税率套利实现长期收益以实现……
1 Constantinides 1983 使用期权理论框架量化税收管理投资中时机损失和收益实现的价值。Wilcox、Horvitz 和 diBartolomeo 2006 对税收管理投资进行了更近期、更全面的阐述。Geddes、Goldberg 和 Bianchi (2015) 说明了指数化损失收获投资组合作为定期产生资本收益的资产配置的一部分的价值。2 Jeffrey 和 Arnott 1993;Arnott、Berkin 和 Ye 2000;以及 Arnott、Berkin 和 Bouchey 2011 证明了税收管理对应税投资者股票投资组合的重要性。Berkin 和 Ye 2003 使用蒙特卡罗模拟量化最高进先出 (HIFO) 会计的收益,以及在股票特定风险相对较高、平均回报率较低和股息收益率较高的市场中损失收获的增量收益。 Berkin 和 Luck 2010 使用蒙特卡罗模拟来展示税收管理在扩展股票策略中的好处。Bergstresser 和 Pontiff 2013 使用实证数据记录了税收对投资组合价值、增长和规模的影响,并表明税收加剧了股票溢价之谜。Israel 和 Moskowitz 2012 探讨了税收管理对规模、价值、增长和动量的影响。Geddes 2011 和 Geddes 和 Tymoczko 2019 使用蒙特卡罗模拟来展示单独管理账户中的损失收割相对于持有交易所交易基金 (ETF) 的税后收益。Sialm 和 Sosner 2018 量化了税收管理的 130-30 和多空市场中性策略的税后回报。 Santodomingo、Nemtchinov 和 Li (2016) 和 Goldberg、Hand 和 Cai (2019a) 使用历史回溯测试研究了流行因子倾向的税后回报和风险状况。Goldberg、Hand 和 Cai (2019b) 描述了指数跟踪和无碳策略的类似概况。3 Constantinides (1984);Dammon、Dunn 和 Spatt (1989);以及 Stein、Vadlamudi 和 Bouchey (2008) 使用蒙特卡洛模拟来评估税率套利策略。

细胞凋亡与癌症关系的系统评价
细胞死亡途径最早由罗伯特·霍维茨在研究秀丽隐杆线虫等低等生物的细胞命运时发现,这最终帮助他获得了 2002 年诺贝尔生理学或医学奖。人们对细胞死亡机制有很多了解,包括从细胞内部和免疫系统 [1] 。其中最重要的一种机制是细胞凋亡,这是一种程序性细胞死亡,可有效清除受损细胞,例如在发育过程中或脱氧核糖核酸 (DNA) 损伤后 [2] 。细胞凋亡的一个重要特征是它从根本上通过一种叫做半胱天冬酶的丝氨酸蛋白酶亚型发挥作用,半胱天冬酶是一种半胱氨酰蛋白酶,可以通过蛋白水解裂解不同的细胞核和细胞质成分。这些胱天蛋白酶由11个成员组成,分为三大组,其中第二组(胱天蛋白酶2、3、7)和第三组(胱天蛋白酶6、8、9、10)参与细胞凋亡。胱天蛋白酶最终依靠不同的信号通路导致细胞的破坏[3]。细胞凋亡的发病机制复杂,涉及两种主要信号通路:外在和内在。两者都会激活效应凋亡胱天蛋白酶,最终导致细胞凋亡特征性的形态和生化改变[4,5]。决定细胞是否凋亡的最重要因素之一是促凋亡和抗凋亡蛋白调节剂之间的平衡。在癌前病变中,DNA损伤可诱导细胞凋亡以清除潜在的有害细胞,从而阻止肿瘤生长。相反,凋亡的紊乱会导致细胞增殖不受控制、癌症发展以及癌症对药物疗法的耐药性 [6]。癌细胞通常会过度表达不同的蛋白质,这些蛋白质在抵抗细胞凋亡的级联反应中起着重要作用。癌细胞诱导的多种机制将它们从程序性细胞死亡中拯救出来,尤其是通过抗凋亡分子的过度表达 [7]。事实上,大多数凋亡信号研究依赖于 B 细胞淋巴瘤 2 同源性 3 (BH3) 蛋白 [8]。促存活和促死亡的 BH3 蛋白之间存在平衡。当这种平衡偏向促死亡的 BH3 蛋白时,往往会发生细胞凋亡,但是当它偏向促存活蛋白时,就会导致存活信号的激活,从而导致癌症等病理状况。

细胞凋亡与癌症关系的系统评价
细胞死亡途径最早由罗伯特·霍维茨在研究秀丽隐杆线虫等低等生物的细胞命运时发现,这最终帮助他获得了 2002 年诺贝尔生理学或医学奖。人们对细胞死亡机制有很多了解,包括从细胞内部和免疫系统 [1] 。其中最重要的一种机制是细胞凋亡,这是一种程序性细胞死亡,可有效清除受损细胞,例如在发育过程中或脱氧核糖核酸 (DNA) 损伤后 [2] 。细胞凋亡的一个重要特征是它从根本上通过一种叫做半胱天冬酶的丝氨酸蛋白酶亚型发挥作用,半胱天冬酶是一种半胱氨酰蛋白酶,可以通过蛋白水解裂解不同的细胞核和细胞质成分。这些胱天蛋白酶由11个成员组成,分为三大组,其中第二组(胱天蛋白酶2、3、7)和第三组(胱天蛋白酶6、8、9、10)参与细胞凋亡。胱天蛋白酶最终依靠不同的信号通路导致细胞的破坏[3]。细胞凋亡的发病机制复杂,涉及两种主要信号通路:外在和内在。两者都会激活效应凋亡胱天蛋白酶,最终导致细胞凋亡特征性的形态和生化改变[4,5]。决定细胞是否凋亡的最重要因素之一是促凋亡和抗凋亡蛋白调节剂之间的平衡。在癌前病变中,DNA损伤可诱导细胞凋亡以清除潜在的有害细胞,从而阻止肿瘤生长。相反,凋亡的紊乱会导致细胞增殖不受控制、癌症发展以及癌症对药物疗法的耐药性 [6]。癌细胞通常会过度表达不同的蛋白质,这些蛋白质在抵抗细胞凋亡的级联反应中起着重要作用。癌细胞诱导的多种机制将它们从程序性细胞死亡中拯救出来,尤其是通过抗凋亡分子的过度表达 [7]。事实上,大多数凋亡信号研究依赖于 B 细胞淋巴瘤 2 同源性 3 (BH3) 蛋白 [8]。促存活和促死亡的 BH3 蛋白之间存在平衡。当这种平衡偏向促死亡的 BH3 蛋白时,往往会发生细胞凋亡,但是当它偏向促存活蛋白时,就会导致存活信号的激活,从而导致癌症等病理状况。

标题“Siri,给我看 AI 的恐怖图像”:基于文本的框架和视觉效果对人工智能支持的影响
人工智能(AI)——定义为模仿人类思维的计算机程序或系统(Russell & Norvig,2003;Turing,1950)——在媒体信息中被描绘成社会进步的工具和充满危险的潘多拉魔盒(Chuan 等人,2019;Obozintsev,2018)。在过去的半个世纪里,《2001:太空漫游》(1968 年)、《终结者》(1984 年)、《黑客帝国》(1999 年)和《我,机器人》(2004 年)等电影都描绘了威胁人类的人工智能(Perkowitz,2007 年)。然而,近年来,人工智能已成为日常生活的一部分,其应用范围广泛,从 Siri 和 Alexa 等虚拟助手到医疗诊断工具。鉴于人工智能日益突出,不仅要了解公众如何看待这项技术,还要了解是什么影响了他们对这项技术的看法,这一点很重要 (Fast & Horvitz, 2017; Neri & Cozman, 2019)。几项全国性调查显示,美国公众对人工智能的看法不一,认为它既有希望,又有潜在的威胁 (Northeastern University & Gallup, 2018; West, 2018; Zhang & Dafoe, 2019)。同样的调查还强调了人口统计、政治倾向和宗教信仰如何预测人们对这项技术的看法。例如,这些调查显示,男性、拥有大学学位的人、年轻人、民主党人和宗教信仰较少的人对人工智能的看法比女性、没有大学学位的人、老年人、共和党人和宗教人士更为积极。相比之下,研究较少关注媒体信息是否(如果是,又是如何影响)对人工智能的看法。然而,媒体对公众对其他新兴技术的看法产生影响的研究结果表明,这些信息可能在促进对人工智能的支持或反对方面发挥重要作用,包括生物技术(Priest,1994)和纳米技术(Brossard 等,2009;Lee & Scheufele,2006)。考虑到这一点,本研究报告借鉴了科学技术的框架和公众舆论理论(Nisbet,2009;Nisbet & Mooney,2007),以探讨不同类型的信息如何影响对人工智能的支持。具体而言,它以先前关于人工智能新闻报道的研究结果为基础(Chuan 等人,2019 年;Obozintsev,2018 年),测试了两种框架对该问题的影响:支持人工智能的“社会进步”框架和反对人工智能的“潘多拉魔盒”框架。该研究还以竞争性框架研究为基础(Chong & Druckman,2007a,2007c;Nisbet 等人,2013 年),通过测试接触竞争性框架对人工智能的影响。同时,它与大多数框架研究(参见 Bolsen 等人,2019 年;Feldman 和 Hart,2018 年;Hart 和 Feldman,2016 年;Powell 等人,2015 年)不同,它研究的是图像(在本例中是现实世界的虚拟助手、现实世界的个人机器人和恐怖电影中的人工智能)如何塑造人们对人工智能的看法,无论是通过图像本身还是与基于文本的框架结合形成的看法。

质疑人工智能预测:影响力驱动的第二意见推荐
[1] Gagan Bansal、Besmira Nushi、Ece Kamar、Dan Weld、Walter Lasecki 和 Eric Horvitz。2019 年。人机协作团队向后兼容的案例。ICML 人机协作学习研讨会 (2019)。[2] Gagan Bansal、Tongshuang Wu、Joyce Zhou、Raymond Fok、Besmira Nushi、Ece Kamar、Marco Tulio Ribeiro 和 Daniel Weld。2021 年。整体是否超过部分?人工智能解释对互补团队绩效的影响。在 CHI 计算机系统人为因素会议论文集上。1-16。 [3] Umang Bhatt、Javier Antorán、Yunfeng Zhang、Q Vera Liao、Prasanna Sattigeri、Riccardo Fogliato、Gabrielle Gauthier Melançon、Ranganath Krishnan、Jason Stanley、Omesh Tickoo 等人。2020 年。不确定性作为透明度的一种形式:测量、传达和使用不确定性。arXiv 预印本 arXiv:2011.07586 (2020)。[4] Zana Buçinca、Maja Barbara Malaya 和 Krzysztof Z Gajos。2021 年。信任还是思考:认知强制函数可以减少人工智能辅助决策对人工智能的过度依赖。ACM 人机交互论文集 5,CSCW1 (2021),1-21。 [5] Rich Caruana、Yin Lou、Johannes Gehrke、Paul Koch、Marc Sturm 和 Noemie Elhadad。2015 年。医疗保健的可理解模型:预测肺炎风险和 30 天内住院率。第 21 届 ACM SIGKDD 国际知识发现和数据挖掘会议论文集。1721–1730。[6] R Dennis Cook。1986 年。局部影响力评估。皇家统计学会杂志:B 系列(方法论)48,2(1986 年),133–155。[7] Maria De-Arteaga、Artur Dubrawski 和 Alexandra Chouldechova。2021 年。利用专家一致性改进算法决策支持。arXiv 预印本 arXiv:2101.09648(2021 年)。 [8] Maria De-Arteaga、Riccardo Fogliato 和 Alexandra Chouldechova。2020 年。《人机交互案例:在存在错误算法评分的情况下做出决策》。2020 年 CHI 计算机系统人为因素会议论文集。1-12。[9] Berkeley J Dietvorst、Joseph P Simmons 和 Cade Massey。2015 年。《算法厌恶:人们在发现算法错误后会错误地避开算法》。《实验心理学杂志:综合》144, 1 (2015),114。[10] Kelwin Fernandes、Jaime S Cardoso 和 Jessica Fernandes。2017 年。《具有部分可观测性的迁移学习应用于宫颈癌筛查》。《伊比利亚模式识别与图像分析会议》。Springer,243-250。 [11] Yarin Gal 和 Zoubin Ghahramani。2016 年。Dropout 作为贝叶斯近似:表示深度学习中的模型不确定性。在国际机器学习会议 (ICML) 中。1050–1059。[12] Ruijiang Gao、Maytal Saar-Tsechansky、Maria De-Arteaga、Ligong Han、Min Kyung Lee 和 Matthew Lease。2021 年。通过 Bandit 反馈实现人机协作。IJCAI (2021)。[13] Pang Wei Koh 和 Percy Liang。2017 年。通过影响函数理解黑盒预测。在第 34 届国际机器学习会议论文集-第 70 卷中。JMLR。org,1885–1894 年。[14] Himabindu Lakkaraju、Jon Kleinberg、Jure Leskovec、Jens Ludwig 和 Sendhil Mullainathan。2017 年。选择性标签问题:在存在不可观测因素的情况下评估算法预测。第 23 届 ACM SIGKDD 国际知识发现与数据挖掘会议论文集。ACM,275–284。