XiaoMi-AI文件搜索系统
World File Search SystemILC2

ILC2 衍生的 LIF 促进从组织免疫到全身免疫的进展
免疫细胞的迁移和归巢对于免疫监视至关重要。运输由粘附和趋化因子受体的组合介导,它们响应趋化因子信号引导免疫细胞到达组织和淋巴系统内的特定位置,以支持组织局部免疫反应和全身免疫 1,2 。我们在这里表明,中断第 2 组先天淋巴细胞 (ILC2) 产生的白血病抑制因子 (LIF) 会阻止免疫细胞离开肺部迁移到淋巴结 (LN)。在没有 LIF 的情况下,病毒感染会导致浆细胞样树突状细胞 (pDC) 滞留在肺部,在那里它们会改善组织局部的抗病毒免疫力,而慢性肺部过敏原攻击会导致明显的免疫细胞积聚和肺部三级淋巴结构的形成。在这两种情况下,免疫细胞都无法迁移到淋巴系统,导致 LN 反应严重受损。从机制上讲,ILC2 衍生的 LIF 诱导肺淋巴管内壁淋巴管内皮细胞产生趋化因子 CCL21,从而允许 CCR7 + 免疫细胞(包括树突状细胞)归巢到淋巴结。因此,ILC2 衍生的 LIF 决定免疫细胞从肺部流出,以调节组织局部免疫与全身免疫以及肺部过敏原和病毒反应之间的平衡。

Mait细胞调节哮喘的潜力
尽管哮喘治疗的最新进展,但仍然需要寻找新的疗法,因为仍然有复发性哮喘患者患者患者对现有疗法的反应不佳。由于第2组先天淋巴样细胞(ILC2)通过触发和加剧2型腹膜弹性在哮喘中起关键作用,因此控制ILC2S功能是对抗严重哮喘的关键。粘膜相关的不变T(MAIT)细胞是人类丰富的先天T细胞,并以T细胞受体依赖性和非依赖性方式激活。mait细胞分别基于转录因子T-bet和ror g t的表达组成。mait细胞在宿主防御病原体和组织修复中起关键作用,对于维持免疫和止血至关重要。我们最近的研究表明,MAIT细胞抑制了ILC2的增殖和气道炎症模型中的功能。mait细胞可以通过促进气道上皮细胞屏障修复和抑制ILC2来减轻两种方式的气道炎症。因此,促进MAIT细胞介导的ILC2增殖和功能抑制的试剂,或设计师MAIT细胞(基因设计以抑制ILC2或促进气道损害的修复),可能是严重哮喘的治疗方法。©2023日本过敏症学会。由Elsevier B.V.这是CC下的开放访问文章(http://creativecommons.org/licenses/4.0/)。
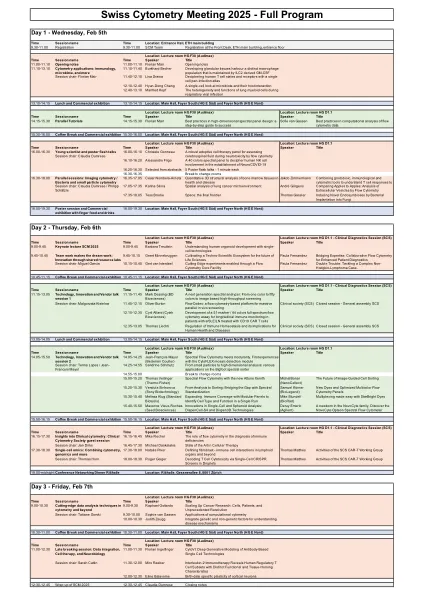
2025瑞士细胞术会议计划草案V3
11.10-11.40 Burkhard Becher Developing glandular tissues harbour a distinct macrophage population that is maintained by ILC2 derived GM-CSF Session chair: Florian Mair 11.40-12.10 Lisa Dratva Deciphering human T cell states and receptors with a single cell pan-infection atlas 12.10-12.40 Hyun-Dong Chang A single-cell look at microbiota and their host interaction 12.40-13.10 Manfred Kopf呼吸病毒感染期间肺髓样细胞的异质性和功能

注射:药物 T 政策
气道炎症是哮喘发病机制中的一个重要组成部分。多种细胞类型(例如肥大细胞、嗜酸性粒细胞、中性粒细胞、巨噬细胞、淋巴细胞、ILC2 细胞)和介质(例如组胺、二十烷酸、白三烯、细胞因子)参与气道炎症。使用 tezepelumab-ekko 阻断 TSLP 可减少与炎症相关的生物标志物和细胞因子,包括血液嗜酸性粒细胞、气道黏膜下嗜酸性粒细胞、IgE、FeNO、IL-5 和 IL-13;然而,tezepelumab-ekko 在哮喘中的作用机制尚未明确。

先天淋巴样细胞 - 免疫的无释放的监护人
先天淋巴样细胞(ILC)表现出显着的可塑性和影响的免疫力,作为致病提示的早期剂量[1-4]。一般术语“ ILC”描述了具有淋巴细胞最常见特征的先天免疫细胞。ilcs包含5种不同的子集:细胞毒性天然杀伤细胞(NK细胞),淋巴组织诱导者细胞(LTI)和3个类似助手的子集,第1组ILCS(ILC1),第2组ILCS(ILC2)(ILC2)和3 ILCS(ILC 3 ILCS)(ILCS(ILC3)[3 ilc3)[3,5-7] [3,5-7]。ILC由它们的特异性细胞因子和转录因子曲线定义,在很大程度上反映了T助手(Th)子集的功能[3,4,8]。然而,ILC的命运是由ID2确定的,ID2阻断了T细胞分化并驱动ILC发育[2,9,10]。因此,ILC通常缺乏抗GEN特异性T或B细胞受体[3],将它们在适应性和先天免疫之间的作用夹在其中。在稳态中,ILC1s依赖于T-bet和Express干扰素γ(IFN-γ)以及肿瘤坏死因子alpha(TNF-α)。ilc2s在分化后与GFI1一起参与GATA3,RORα,BCL11B [2,3],并产生IL-13,IL-5,IL-4,IL-4,IL-9和两静脉蛋白[2,11,12]。最后,在分泌IL-22和IL-17时,ILC3需要RORγT以及AHR [4,13 - 15]。大多数ILC是长期寿命的自我更新细胞,居住在组织屏障上,如果他们通过细胞因子释放很容易对生理或病理触发反应[2,4,16,17](表1)。因此,ILC可能获得与有效免疫监测有关的持久免疫记忆[18-21]。我们将讨论针对不同病原体的可塑性。ilcs还可以为引起慢性发病的免疫反应失调,从而恶化疾病结果[4,22,23](表1)。然而,调节ILC对病原体极化的生物学机制仍然不足,对NK细胞和ILC之间的可塑性的了解甚至更少[4,24]。此外,人类和鼠ILC之间的相当大差异一直在摆动路障,以更好地了解其免疫学功能[25,26]。在这项工作中,我们提供了有关主要ILC子集的概述的概述,强调了在传染病中的可能作用。最后,我们将检查ILC在患者中的免疫调节潜力,以及ILC生物学的未来挑战。

IL-2恢复免疫力为不同的细胞电路
白介素2(IL-2)是关键的稳态细胞因子,在免疫原性和耐受性免疫调节中都具有治疗性应用。临床用途受到多效性功能和广泛的受体表达的阻碍,并具有意外的不良事件。在这里,我们开发了一种新型的小鼠菌株来转移IL-2的产生,从而识别上下文结果。网络分析确定了Treg和Treg和常规CD4 T细胞中IL-2产生的竞争性适应性成本的优先访问。CD8 T和NK细胞表现出对自分泌IL-2产生的偏爱。IL-2来自树突状细胞扩增的Tregs,而B细胞产生的IL-2诱导了两个依赖于上下文的电路:CD8 + Tregs和ILC2细胞的急剧扩张,后者驱动下游下游,IL-5 - 介导的,eried,嗜酸性粒细胞电路。源特异性效应证明了IL-2功能的上下文影响,并有可能解释在临床试验期间观察到的不良影响。靶向的IL-2产生有可能基于临床可取性在IL-2网络中放大或淬灭特定电路。

供体淋巴细胞输注和细胞疗法
结果:在基线时确定了十二个基于其转录谱的角膜免疫细胞群体,由单核细胞,居民(RMP)和MMP12/13高巨噬细胞,树突状细胞(CDC2),中性粒细胞,中性粒细胞,肥大细胞,T/B细胞,PER T/B细胞和天生(γdtt和gudinte nk and l canty)和nk和l lcc2 and y lcc2 and lcc2和l lcc2 and lcc2 and y l l l l l l l l l l l l l l l l l l l l l l canty and contion nk and l canty and。T细胞和常驻巨噬细胞(RMP)分别是正常角膜中最大的人群,分别占18.6%和18.2%。rmp增加到55.2%的细胞。随着细胞因子和趋化因子(TNF,CXCL1,CCL12,IL1RN)的增加,RMP中的1,365个基因表达显着变化(adj p <0.0001),炎症标记(VCAM,ADAM17,JUNB),TAM受体(TAM受体(MERTK)和SEROME和SENES和M.HC和M.HC和MHC。发现了从单核细胞到末端状态RMP的分化轨迹。吞噬作用,C型凝集素受体信号传导,NF-kappa B信号传导和Toll样受体信号传导是这些细胞活性增强的途径之一。MRC1 + RMP的百分比在角膜中增加,并且在与上皮神经丛相邻的基底上皮中观察到它们。趋化因子CXCL1的浓度在角膜中增加,并增加了对局部施用的高渗盐水的刺激/疼痛反应。

缺氧诱导因子2α促进保护性TH2细胞...
Hypoxia-inducible factor 2 α promotes protective Th2 cell responses during intestinal 1 helminth infection 2 3 Jasmine C. Labuda 1 , Tayla M. Olsen 1,2 , Sheenam Verma 1 , Samantha Kimmel 1 , Thomas H. 4 Edwards 3 , Matthew J. Dufort 3 , Oliver J. Harrison 1,4 5 6 1 Center for Fundamental Immunology, Benaroya Research美国华盛顿州西雅图研究所。7 2分子和蜂窝生物学计划,美国华盛顿州西雅图市华盛顿大学。8 3美国华盛顿州西雅图市贝纳罗亚研究所系统免疫学中心。9 4美国华盛顿州华盛顿大学华盛顿大学免疫学系。10 11通信:oharrison@benaroyaresearch.org 12 13摘要:TH2细胞必须感知并适应组织环境,以提供保护性宿主14免疫和组织修复。在这里,我们检查了促进Th2细胞15分化和功能的机制。单细胞RNA-seq 16分析来自小肠道椎板椎板的CD4 + T细胞17揭示了基因EPAS1的高表达,编码了转录因子缺氧缺氧诱导的18因子2a(HIF2α)。在体外,即使在非极化条件下,暴露于缺氧或遗传HIF2α激活也促进了Th2细胞19分化。在小鼠中,CD4 + T细胞中的HIF2α激活20在没有感染的情况下促进了肠道Th2细胞的积累,而HIF2α缺陷21受损的CD4 + T细胞介导的宿主对肠舵感感染的免疫免疫。24 25简介:肠蠕虫感染是全球最普遍的慢性感染26。我们的发现22确定了缺氧,氧调节的转录因子缺氧诱导因子2α23(HIF2α)是小肠内Th2细胞分化和功能的关键调节剂。Helminth infections are often associated with polarized “type 2” immunity, including 27 activation and accumulation of T helper 2 (Th2) cells, type-2 innate lymphoid cells (ILC2), tissue 28 basophils and eosinophils, elevated serum immunoglobulin E (IgE), alternative activation of 29 macrophages and alterations of epithelial differentiation and mucus production that统称30重塑感染的解剖部位2。免疫事件和31个组织重塑的类似级联反应引发局部组织病理学发生在过敏性疾病中,包括过敏32哮喘3。33 34指导屏障组织中Th2细胞分化的机制尚不清楚。35然而,证据支持组织微环境在建立36保护性Th2细胞分化和功能中的指导性作用,这是由染色质访问性37和/或基因表达的变化提供的,在将Th2细胞从淋巴结到本塞质体38组织4,5的TH2细胞转运后的基因表达。组织警报蛋白,包括IL-25,IL-33和TSLP是在39个屏障组织中产生的关键因素,这些因素在Helminth 40感染6,7期间共同促进2型免疫力和Th2细胞反应。在41个屏障组织中影响Th2细胞功能的组织环境中其他提示的身份仍有待鉴定。42 43缺氧诱导因子(HIF)是介导细胞的关键转录因子,对缺氧8的有机体反应4。Consisting of 3 family members, (HIF1 α , HIF2 α and HIF3 α , 45 encoded by Hif1a , Epas1 and Hif3a, respectively), HIFs are post-translationally modified in an 46 oxygen-dependent enzymatic cascade that regulates their stability, nuclear translocation, 47 binding to hypoxia-response elements (HRE) and transcription of低氧诱导基因8。48在常氧条件下,HIF蛋白通过氧气在关键的脯氨酸残基上通过氧气-49依赖性丙酰羟化酶(PHD)酶羟基氧化。通过50