XiaoMi-AI文件搜索系统
World File Search SystemISGS


干扰素刺激基因的过度表达筛选确定 RARRES3 是弓形虫感染的限制因子
摘要 弓形虫是一种重要的人类病原体,全球约有三分之一的人感染该病。细胞因子干扰素γ (IFNγ) 在感染过程中被诱导,对于限制人体细胞中的弓形虫生长至关重要。生长限制被认为是由于干扰素刺激基因 (ISG) 的诱导所致,这些基因被上调以保护宿主免受感染。尽管 IFNγ 可诱导数百种 ISG,但它们在限制人体细胞中寄生虫生长方面各自的作用仍然有些难以捉摸。为了解决这一缺陷,我们筛选了 414 个 IFNγ 诱导的 ISG 库,以确定影响人体细胞中弓形虫感染的因子。除了可能通过诱导大量下游基因发挥作用的 IRF1 之外,我们还发现 RARRES3 是限制多种人体细胞系中弓形虫过早逸出而产生的单一因子。总体而言,虽然我们成功鉴定出一种新的 IFNγ 诱导的可限制弓形虫感染的因子,但单独表达时能够限制弓形虫感染的 ISG 数量有限,这表明 IFNγ 介导的对弓形虫感染的免疫是一个复杂的、多因素的过程。

海德堡材料PowerPoint Master
•Prime = ISGS,Heidelberg Materials =主机和工业合作伙伴,Drilling = Projeo,以及较小的团队成员,包括:Indiana Geological&Water Survey,Trimeric和Gnarly Tree Tree Sustainability Institute

内源性逆转录病毒表达在间皮瘤发育的实验小鼠模型中激活I型干扰素信号
在间皮瘤发育实验模型中,早期事件包括双链RNA(DSRNA)中编辑水平的增加。我们假设内源性逆转录病毒(ERV)的表达有助于DSRNA形成和I型干扰素信号传导。与非肿瘤样品相比,肿瘤的 ERV和干扰素刺激的基因(ISG)表达明显更高。 12个肿瘤特异性ERV(“ Mesoerv1-12”)被鉴定出来并通过qPCR在小鼠组织中验证。 与间皮瘤细胞相比,小鼠胚胎成纤维细胞(MEF)的“ Mesoerv1-12”表达较低。 “ Mesoerv1-12”水平通过脱甲基化剂5-Aza-2' - 脱氧胞苷的处理显着提高,并伴随着DSRNA和ISGS的水平升高。 与MEF相比,间皮瘤细胞中的基底ISGS表达更高,并且通过阻断IFNAR1和沉默的MAVS,JAK抑制剂r梭替尼显着降低了。 “ Mesoerv7”启动子在5-Aza-CDR处理后,与假小鼠组织以及间皮瘤细胞以及MEF细胞和MEF相比,在石棉暴露的暴露中被脱甲基化。 这些观察结果发现了石棉诱导的间皮瘤的新颖方面,从而导致ERV表达因启动子去甲基化而引起,并且与DSRNA水平的增加和IFN型信号传导的激活相似。 这些特征对于早期诊断和治疗很重要。ERV和干扰素刺激的基因(ISG)表达明显更高。12个肿瘤特异性ERV(“ Mesoerv1-12”)被鉴定出来并通过qPCR在小鼠组织中验证。与间皮瘤细胞相比,小鼠胚胎成纤维细胞(MEF)的“ Mesoerv1-12”表达较低。“ Mesoerv1-12”水平通过脱甲基化剂5-Aza-2' - 脱氧胞苷的处理显着提高,并伴随着DSRNA和ISGS的水平升高。与MEF相比,间皮瘤细胞中的基底ISGS表达更高,并且通过阻断IFNAR1和沉默的MAVS,JAK抑制剂r梭替尼显着降低了。“ Mesoerv7”启动子在5-Aza-CDR处理后,与假小鼠组织以及间皮瘤细胞以及MEF细胞和MEF相比,在石棉暴露的暴露中被脱甲基化。这些观察结果发现了石棉诱导的间皮瘤的新颖方面,从而导致ERV表达因启动子去甲基化而引起,并且与DSRNA水平的增加和IFN型信号传导的激活相似。这些特征对于早期诊断和治疗很重要。

酪氨酸激酶抑制剂耐药的克隆进化
中东呼吸道综合征冠状病毒(MERS-COV)感染会导致人类致命的肺部炎症性疾病。相反,骆驼和蝙蝠是主要的储层宿主,耐受的MERS-COV复制而不患有临床疾病。在这里,我们从MERS-COV康复的骆驼中分离了宫颈淋巴结(LN)细胞,并用两种不同的病毒菌株(进化枝B和C)脉冲它们。病毒复制,但安装了细胞免疫反应。让人联想的Th1反应(IFN-G,IL-2,IL-12),并伴随着抗病毒反应的明显且短暂的峰值(I型IFNS,IFNS,IFN-L 3,ISGS,ISGS,PRRS和TFS)。重要的是,炎症细胞因子(TNF-A,IL-1 B,IL-6,IL-8)的表达或膨胀成分(NLRP3,CASP1,Pycard)的表达被抑制。讨论了IFN-L 3在骆驼物种中对平衡量弹性过程以及桥接先天和适应性免疫反应的作用。我们的发现阐明了有关在没有临床疾病的情况下如何控制MERS-COV的关键机制。

RNA 编辑在有限数量的位点足以阻止小鼠大脑中的 MDA5 激活
作用于 RNA 1 的腺苷脱氨酶 (ADAR1) 是一种负责腺苷到肌苷 RNA 编辑的酶,由两种亚型组成:核 p110 和细胞质 p150。小鼠中 Adar1 或 Adar1 p150 基因的缺失会导致胚胎致死,并伴有干扰素刺激基因 (ISG) 的过表达,这是由于黑色素瘤分化相关蛋白 5 (MDA5) 对未编辑的内源转录本的异常识别所致。然而,在众多 RNA 编辑位点中,有多少 RNA 位点需要编辑,尤其是由 ADAR1 p150 编辑,以避免 MDA5 激活,以及 ADAR1 p110 是否有助于此功能仍不清楚。具体来说,ADAR1 p110 在小鼠脑中含量丰富,而 ADAR1 p150 的表达量微乎其微,而 ADAR1 突变会导致艾卡迪-痛风综合征,在这种综合征中,大脑是受影响最严重的器官之一,同时伴有 ISG 表达升高。因此,了解 RNA 编辑介导的预防大脑中 MDA5 活化的方法尤为重要。在这里,我们建立了 Adar1 p110 特异性敲除小鼠,在这种小鼠中未观察到 ISG 表达上调。这一结果表明 ADAR1 p150 介导的 RNA 编辑足以抑制 MDA5 活化。因此,我们进一步创建了 Adar1 p110 / Adar2 双敲除小鼠来确定 ADAR1 p150 介导的编辑位点。这项分析表明,尽管没有观察到 ISG 表达升高,但在 Adar1 p110 / Adar2 双敲除小鼠的大脑中,只有不到 2% 的编辑位点得以保留。值得注意的是,我们发现一些位点被高度编辑,与野生型小鼠的编辑位点相当,这表明存在 ADAR1 p150 特异性位点。这些数据表明,在非常有限的位点上进行 RNA 编辑(由少量 ADAR1 p150 介导)足以阻止 MDA5 激活,至少在小鼠大脑中是如此。

转座元件的压抑增强了干扰素beta信号传导和
图1:与光子相比,质子辐射的类器官显示出更高的自我更新能力和IFN-β响应。(a)自我更新测定法的示意图。在第5天进行辐照,并在第18天的自我更新后计算了按器官形成效率(Ofe%)的定量。(b)自我更新后培养中器官的代表性图像。比例尺,100 µm。(c)相对于对照样品的折叠变化(FC)所示的器官定量。n = 9动物/病情。(d)大量RNA-Seq分析的示意图。RNA。(e)前10名重要(p.adj。<0.05)在辐照后2和6天,质子与控制和光子对照中的生物过程。(f)显着(p <0.05)ISG的基因表达水平从辐照后6天的类器官的大量RNA-seq数据推断出来。数据相对于对照样品显示为log 2 FC。(g)辐照后6天对ISG的RT-QPCR分析。数据相对于对照样品显示为FC。n = 4个动物/状况。(h)辐照后6天后对类器官的STAT1,PSTAT1和GAPDH的Western印迹分析。(i)STAT1的蛋白质印迹定量(图1i和S2e)。STAT1蛋白水平的GAPDH标准化。 数据相对于对照样品显示为FC。 n = 6只动物/状况。 数据是平均值±s.e.m。 学生的T检验和双向ANOVA。 *p <0.05,** p <0.01。STAT1蛋白水平的GAPDH标准化。数据相对于对照样品显示为FC。n = 6只动物/状况。数据是平均值±s.e.m。学生的T检验和双向ANOVA。*p <0.05,** p <0.01。另请参见图S1和S2。
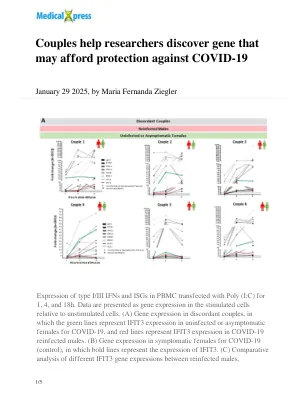
夫妇帮助研究人员发现可能能够保护Covid-19
I/III型IFN和ISG在PBMC中用Poly(I:C)转染为1、4和18H的表达。数据相对于未刺激的细胞,将数据作为基因表达表示。(a)不一致的夫妻中的基因表达,其中绿线代表未感染或无症状的女性在covid-19中表示IFIT3的表达,而红线表示在Covid-19的COVID-19再感染的雄性中的IFIT3表达。(b)在症状女性中的基因表达于19(对照),其中粗线表示IFIT3的表达。(c)对重新感染的雄性之间不同IFIT3基因表达的比较分析,

实施基于市场的经济调度(MBED)
安全约束经济调度及其与 MBED 的主要区别 MBED 概念的扩展是安全约束经济调度 (SCED),由 POSOCO 从 2019 年 4 月开始实施,最初使用一组较小的发电机。SCED 在制定最终计划并在实时市场 (RTM) 关闭后,优化了主要州际发电站 (ISGS) 站(以及其他参与的 IPP 和州发电公司)的电力生产成本。CERC 已将 SCED 的实施延长至 2021 年 9 月。优化的主要手段是按优先顺序重新安排计划发电机的调度,这样,在技术限制下,先将运营成本较低的发电机全部预订,然后再调度运营成本较高的发电机。
将抗病毒药物与 BET 抑制剂联合使用有利于对抗 SARS‐CoV‐2 感染
和对 SARS-CoV-2 生命周期至关重要的病毒蛋白,并证明了溴结构域和末端外结构域蛋白 2 和 4 (BRD2/BRD4) 与 SARS-CoV-2 的 E 蛋白相互作用。另一项利用 CRISPRi 筛选的研究表明,BRD2 抑制会下调血管紧张素转换酶 2 (ACE2) 表达,并通过下调干扰素刺激基因 (ISG) 来控制 COVID-19 患者的过度活跃免疫反应。6 此外,Gilham 等人报道,溴结构域和末端外结构域蛋白家族 (BET) 抑制剂阿帕贝龙 (RVX-208) 通过降低 ACE2 表达来阻断 SARS-CoV-2 感染,7 而 Qiao 等人表明,使用 BET 抑制剂对雄激素受体的转录抑制也会导致 SARS-CoV-2 感染减少。8