XiaoMi-AI文件搜索系统
World File Search SystemISGS

表征病理相关的人类肺上皮细胞中针对 SARS-CoV-1 和 -2 感染的时间和整体宿主先天免疫反应
严重急性呼吸综合征冠状病毒 1 (SARS-CoV-1) 和 2 (SARS-CoV-2) 是已在世界范围内造成大量发病率和死亡率的 β 冠状病毒 (β-CoV)。因此,更好地了解宿主对 β-CoV 的反应将有助于深入了解这些病毒的发病机制,从而确定医疗对策的潜在目标。在本研究中,我们的目标是使用系统生物学方法探索 SARS-CoV-1 和 -2 感染在病理相关的人肺上皮细胞 (Calu-3/2B4 细胞) 中随时间引发的先天免疫反应的规模和范围。在 β-CoV 或模拟感染 Calu-3/2B4 细胞后 12、24 和 48 小时提取的总 RNA 进行 RNA 测序和功能富集分析,以选择感染后表达显着调节的基因。结果表明,SARS-CoV-1 和 -2 在病理相关的人肺上皮细胞中刺激了相似但不同的先天抗病毒信号通路。此外,我们发现许多与病毒生命周期、干扰素和干扰素刺激基因 (ISG) 相关的基因在多个时间点上调。基于它们在被 SARS-CoV-1、SARS-CoV-2 和 Omicron BA.1 感染后的显著调节,四种 ISG,即骨髓基质细胞抗原 2 ( BST2 )、Z-DNA 结合蛋白 1 ( ZBP1 )、CXC 基序趋化因子配体 11 ( CXCL11 ) 和干扰素诱导的跨膜蛋白 1 ( IFITM1 ),被确定为针对 β -CoV 的潜在药物靶点。我们的研究结果表明,这些基因通过先天免疫反应直接或间接影响病原体,使其成为宿主定向抗病毒药物的潜在靶点。总之,我们的结果表明,SARS-CoV- 1 和 SARS-CoV-2 感染对宿主先天免疫反应产生不同的影响。

首次充电/通电(FTC)和...
d。 “受益人”是指在州际生成站中享有股份的人; e。 “委员会”是指该法第76条第(1)款中提到的中央电力监管委员会; f。卖方时块中的“偏差”是指其实际注入总额减去其计划的总生成,而对于买方来说,其实际的实际抽奖减去了其总计划的抽奖; g。 “干扰记录器(DR)”是指在事件期间提供系统参数的预选数字和模拟值的行为; h。 “事件记录设施”是指提供的设备来记录继电器和其他设备的按时间顺序排列的操作序列;我。“网格代码”是指委员会根据该法第79条第(1)款规定的网格代码; j。“州际生成station(ISGS)”是指中央生成站或其他生成站,其中两个或多个州具有股票;

核DHX9与STAT1合作以转录干扰素刺激的基因
RNA解旋酶DHX9已被广泛地描述为转录调节剂,这与其主要是核定位置一致。它也参与了识别细胞质中的RNA病毒。但是,没有体内数据来支持DHX9的抗病毒作用。同时,作为一种核蛋白,IF以及核DHX9如何促进抗病毒免疫仍然在很大程度上未知。在这里,我们产生了髓样特异性和肝细胞 - 特异性DHX9敲除小鼠,并确认DHX9对于体内RNA病毒感染的宿主抗性至关重要。通过DHX9缺乏小鼠的其他基因敲除MAV或STAT1,我们证明了核DHX9在调节I型干扰素下游的干扰素刺激的基因(ISG)表达中起着积极的作用。Mech-在干扰素刺激下,DHX9直接与STAT1结合,并将POL II招募到ISG启动子区域,以参与ISGS的STAT1介导的转录。共同发现了核DHX9在抗病毒免疫中的重要作用。

MX2介导的对HIV-1的先天免疫受到丝氨酸磷酸化的调节
抗病毒细胞因子干扰素(IFN)激活IFN刺激基因(ISGS)的表达以建立抗病毒态。粘菌病毒抗性2(MX2/MXB)是一种ISG,它抑制了HIV-1的核进口并与病毒式衣壳和细胞核转运机械相互作用。我们将肌球蛋白轻链磷酸酶(MLCP)亚基MyPT1和PPP1CB作为MX2的正常作用调节剂,与其N末端结构域(NTD)相互作用。我们证明了NTD在14、17和18的位置的丝氨酸磷酸化抑制了MX2抗病毒功能,可防止与HIV-1帽骨和核转运因子的相互作用,并由MLCP逆转。重要的是,NTD丝氨酸磷酸化还阻碍了MX2介导的细胞核货物进口的抑制作用。我们还发现,IFN治疗降低了这些丝氨酸处的磷酸化水平,并概述了稳态调节机制,其中通过磷酸化对MX2的抑制以及MLCP介导的去磷酸化的抑制作用,平衡MX2对MX2对正常细胞与HISATE免疫功能的有害作用平衡,与HIV-1抗HIV-1。

第56届中西部学生生物医学研究论坛
背景,意义和假设:沙眼衣原体(CTR)是一种强制性细胞内病原体,是细菌性传播感染(STI)的主要原因。尽管通常无症状,但感染可能会发展为上等生殖道,并可能导致严重的生殖健康后遗症,例如骨盆炎性疾病,异位妊娠,甚至不育(如果未经治疗)。全国有160万例案件,直接终身费用超过6.9亿美元,CTR被认为是主要的公共卫生负担。一些人自发清除感染,这些感染归因于宿主的适应性免疫。然而,研究还表明,感染可能会持续存在,并重新感染表明长期保护性免疫充其量是部分部分。尽管对CTR感染的适应性免疫反应已充分表征,但主动感染如何影响宿主的先天免疫力,尤其是在CTR-上皮界面上仍未开发。此外,通常会忽略存在未感染的旁观者细胞的宫颈上皮感染期间宿主反应的表征,而不是使用完全感染的上皮单层感染模型,其中在单个时间点感染后在单个时间点收集样品。这进一步强调了在感染过程中调查跨多个时间点的宿主反应的需求,这可能对CTR存活和扩散有影响。宫颈上皮位于CTR -HOST相互作用的中心,因为它是感染的主要部位。可溶性因子在内,包括干扰素(IFN)在感染微环境中产生丰富的。上皮相关的IFN(例如IFNβ和IFNλ)以自分泌和旁分泌方式通过JAK-STAT途径驱动IFN刺激的基因(ISGS)的表达。由于大多数ISG是推定的抗菌剂,因此累积上皮反应通常是抗菌剂,有助于病原体限制。因此,感染上皮细胞中的细胞因子信号传导通常是颠覆病原体的目标。我们最近表明,CTR可以抑制完全感染的上皮单层的上皮IFN反应。这与该领域所谓的CTR上皮宿主免疫生物学相反,通常被视为促炎性。为了调和观察到的减毒上皮IFN与当前炎症性CTR上皮相互作用的概念,我们假设旁观者细胞对于塑造细胞因子环境至关重要,并且在塑造上皮IFN反应的失调可能对CTRENSINCAINS产生导致的影响。

微管活性调节激酶 2 增强宿主……
先天免疫系统对于抵御病原体入侵、有效控制感染以及触发适应性免疫反应以消除传染源至关重要。本研究揭示了微管亲和力调节激酶 2 (MARK2) 作为广谱抗病毒免疫调节剂的关键作用,具体通过其与鸟嘌呤核苷酸交换因子 H1 (GEF- H1) 的相互作用以及与 TANK 结合激酶 1 (TBK1) 的结合。至关重要的是,MARK2 的抗病毒功效取决于其激酶活性,特别是其在丝氨酸 645 位点磷酸化 GEF-H1 的能力。该磷酸化事件是激活 TBK1 的关键触发因素,从而导致诱导 I 型干扰素 (IFN-I) 和干扰素刺激基因 (ISG)。我们的结果表明,GEF-H1 是一种 ISG,并由 MARK2 促进。这些发现不仅证实了 MARK2 是 GEF-H1 的激酶,还揭示了 MARK2 增强宿主抗病毒防御的一种以前未被认识的机制。通过对 GEF-H1 进行策略性磷酸化来增强 IFN-I 信号,MARK2 显著增强了抗病毒免疫反应,为细胞防御机制的分子协调提供了新的见解。

CD14+单核细胞衍生的氧化线粒体的作用...
定义有助于病理干扰素(IFN)1型少年性皮肌炎(JDM)的宿主机制的抽象目标。方法在CD4 +,CD8 +,CD14 +和CD19 +细胞上进行了RNA序列,这些细胞从预处理和治疗JDM(预处理n = 10,治疗n = 11)和年龄/性别匹配的儿童健康对照(CHC n = 4)。通过荧光显微镜,通过13 C葡萄糖摄取测定法和氧化的线粒体DNA(OXMTDNA)含量评估线粒体形态和超氧化物,通过dot-blot评估。健康控制PBMC和JDM预处理PBMC与IFN-α,OxmtDNA,CGAS抑制剂,TLR-9拮抗剂和/或N-乙酰半胱氨酸(NAC)培养。通过qPCR测量IFN刺激的基因(ISGS)表达。 功能实验的患者总数和对照组,JDM n = 82,总CHC n = 35。 结果与JDM CD14+单核细胞中ISG表达增加相关的线粒体相关基因表达失调。 线粒体相关基因表达的改变与线粒体生物学的改变相似,包括“巨胶囊成分”,细胞代谢和超氧化物歧化酶(SOD)1的基因表达降低。 这与氧化的线粒体(OXMT)DNA的产生增强有关。 oxmtDNA在健康的PBMC中诱导的ISG表达,通过靶向氧化应激和细胞内核酸感觉途径来阻止。 结论这些结果描述了一种新的途径,其中JDM CD14+单核细胞中的线粒体生物学改变导致OxmtDNA产生并刺激ISG表达。通过qPCR测量IFN刺激的基因(ISGS)表达。功能实验的患者总数和对照组,JDM n = 82,总CHC n = 35。结果与JDM CD14+单核细胞中ISG表达增加相关的线粒体相关基因表达失调。线粒体相关基因表达的改变与线粒体生物学的改变相似,包括“巨胶囊成分”,细胞代谢和超氧化物歧化酶(SOD)1的基因表达降低。这与氧化的线粒体(OXMT)DNA的产生增强有关。oxmtDNA在健康的PBMC中诱导的ISG表达,通过靶向氧化应激和细胞内核酸感觉途径来阻止。结论这些结果描述了一种新的途径,其中JDM CD14+单核细胞中的线粒体生物学改变导致OxmtDNA产生并刺激ISG表达。互补实验表明,在体外实验条件下,通过抗氧化剂NAC,TLR9拮抗剂和较小程度的CGAS抑制剂靶向这些途径,抑制了预处理JDM PBMC中的ISG表达。针对此途径具有JDM和其他IFN 1型自身免疫性疾病的治疗潜力。

免疫原性死亡在癌症免疫疗法中的新兴作用
癌症免疫疗法,例如免疫检查点阻滞(ICB),已成为有效癌症治疗的开创性方法。尽管具有很大的潜力,但临床研究表明,当前对癌症免疫疗法的反应率是次优的,主要归因于某些类型的恶性肿瘤的免疫原性低。免疫原性细胞死亡(ICD)代表一种能够增强肿瘤免疫原性并激活免疫能力宿主中肿瘤特异性和适应性免疫反应的调节细胞死亡(RCD)的形式。因此,对ICD及其进化有更深入的了解对于制定更有效的癌症治疗策略至关重要。本综述仅关注与ICD模式及其机械见解有关的历史和最新发现,尤其是在癌症免疫疗法的背景下。我们最近的发现也被突出显示,揭示了在多动型I型IFN信号传导过程中,非典型干扰素(IFN)刺激基因(ISGS)促进了一种ICD诱导模式,包括Polo样激酶2(PLK2)。审查通过讨论ICD的治疗潜力结束,并特别注意其在癌症免疫疗法领域内的临床前和临床环境中的相关性。
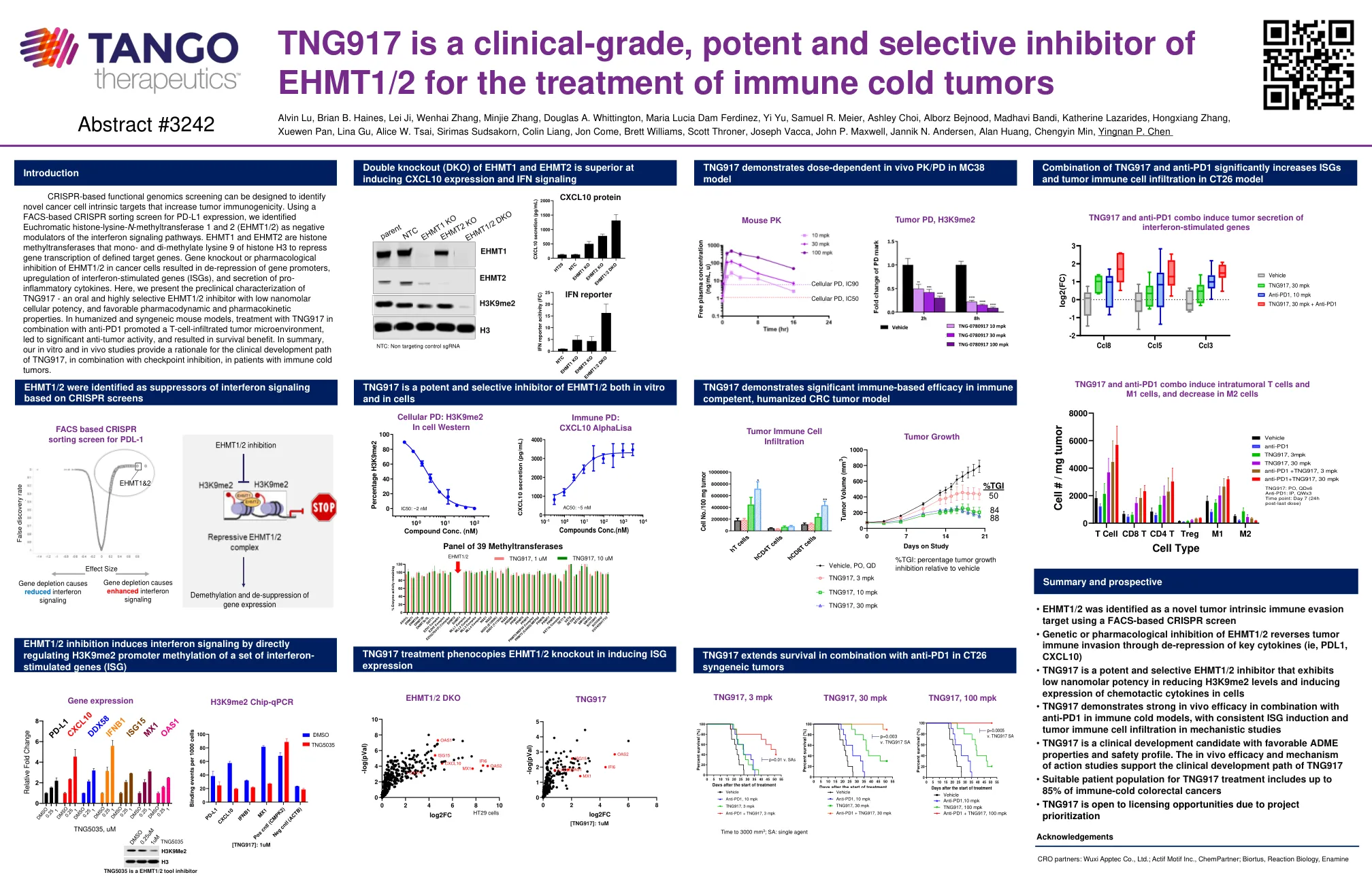
TNG917是一种用于治疗免疫冷肿瘤的临床级,有效和选择性抑制剂
基于CRISPR的功能基因组学筛查可以设计用于鉴定增加肿瘤免疫原性的新型癌细胞固有靶标。使用基于FACS的CRISPR分类屏幕用于PD-L1表达,我们确定了正式的组蛋白 - 赖氨酸-N-甲基转移酶1和2(EHMT1/2)是干扰素信号通路的负调节剂。EHMT1和EHMT2是组蛋白H3的单甲基赖氨酸9的组蛋白甲基转移酶,以抑制定义靶基因的基因转录。基因敲除或对癌细胞中EHMT1/2的药理抑制作用导致基因启动子的抑制,干扰素刺激的基因(ISGS)的上调以及炎性细胞因子的分泌。在这里,我们介绍了TNG917的临床前表征 - 口服和高度选择性的EHMT1/2抑制剂,具有低纳摩尔细胞效力,以及有利的药效和药物性特性。在人性化和合成小鼠模型中,与抗PD1结合使用TNG917处理促进了T细胞浸润的肿瘤微环境,导致了显着的抗肿瘤活性,并带来了生存益处。总而言之,我们的体外和体内研究为免疫冷肿瘤患者的临床发展路径提供了临床发育路径的基本原理。

NOG1通过靶向磷酸化的干扰素调节因子3
先天免疫系统是宿主防御的第一线,研究干扰素(IFN)信号负调控的机制对于维持先天免疫反应的平衡很重要。在这里,我们发现宿主GTP结合蛋白4(NOG1)是先天免疫反应的负调节剂。NOG1的过表达抑制了病毒RNA和DNA介导的信号通路,NOG1缺乏症促进了抗病毒先天免疫反应,从而导致NOG1促进病毒复制的能力。囊泡口腔炎病毒(VSV)和单纯疱疹病毒1型(HSV-1)感染诱导NOG1缺乏小鼠的IFN-β蛋白较高水平。同时,NOG1缺陷型小鼠对VSV和HSV-1感染更具抗性。NOG1通过靶向IRF3抑制I型IFN产生。nog1与磷酸化的IFN调节因子3(IRF3)相互作用,以损害其DNA结合活性,从而下调IFN-β和下游IFN刺激基因(ISGS)的转录。NOG1的GTP结合域是负责此过程的原因。总而言之,我们的研究揭示了NOG1如何通过靶向IRF3对IFN-β进行负面影响的潜在机制,该机制发现NOG1在宿主先天免疫中的新作用。