XiaoMi-AI文件搜索系统
World File Search SystemInhibitor

与免疫检查点抑制剂和 BRAF/MEK 抑制剂治疗相关的腹泻和结肠炎
腹泻和结肠炎是使用免疫检查点抑制剂 (ICI) 的常见不良事件 (AE),包括细胞毒性 T 淋巴细胞相关蛋白 4 (CTLA-4)、程序性细胞死亡蛋白 1 (PD-1) 和程序性死亡配体 1 (PD-L1) 抑制剂 [1-3]。靶向疗法,例如丝氨酸/苏氨酸蛋白激酶 B-Raf (BRAF) 抑制剂和选择性丝裂原活化蛋白激酶激酶 (MEK),已用于治疗具有激活丝裂原活化蛋白激酶途径突变的特定癌症患者 [4-7]。美国食品药品监督管理局已批准多种 BRAF 和 MEK 抑制剂组合用于治疗具有 BRAF 突变的黑色素瘤、肺癌、结肠癌或甲状腺癌患者 [6-7]。在临床试验中,胃肠道不良反应(包括腹泻、腹痛、恶心和呕吐)很常见,并且联合使用 BRAF 和 MEK 抑制剂的患者比单独使用其中一种药物的患者更严重 [6-8]。BRAF 和 MEK 抑制剂诱发胃肠道炎症的机制

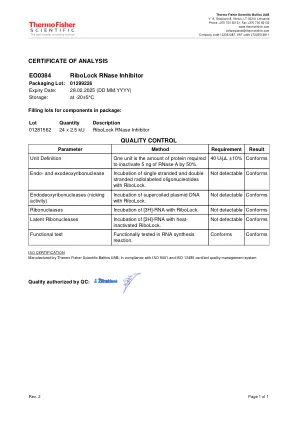
EO0384 RiboLock RNase 抑制剂
ISO 认证由 Thermo Fisher Scientific Baltics UAB 制造,符合 ISO 9001 和 ISO 13485 认证的质量管理体系。

社会支持作为技术压力的抑制剂
摘要 由于数字化的不断发展以及伴随新冠疫情而来的社交距离措施,许多人的工作条件和环境发生了变化。由于远程办公的增多,工作中数字技术用于交流和协作的使用也得到了加强,这可能会导致技术压力。本文利用两项调查的纵向数据(一项在新冠疫情之前,一项在疫情期间),分析了四个社会支持维度(上司支持、同事支持、工作社区意识和家庭支持)与技术压力制造者之间的关系。研究表明,社会支持可以有效抑制技术压力制造者。然而,在这方面,社会支持维度必须有所区别。此外,研究结果还表明,家庭支持的抑制作用在新冠疫情期间变得更加重要。这些结果有助于技术压力研究以及疫情后工作新常态的研究。

Mash中的FASN抑制剂denifanstat
1阶段1 IST博士伊丽莎白公园。 1 Abdul和Al。,Hepatology,2020年7月。 doi:10.1002/没有。 3 3 Loomba and Al。,胃肠病学和肝,9,1090 - 1100。1阶段1 IST博士伊丽莎白公园。1 Abdul和Al。,Hepatology,2020年7月。 doi:10.1002/没有。 3 3 Loomba and Al。,胃肠病学和肝,9,1090 - 1100。1 Abdul和Al。,Hepatology,2020年7月。 doi:10.1002/没有。3 3 Loomba and Al。,胃肠病学和肝,9,1090 - 1100。

Sonrotoclax (BGB-11417) BCL2 抑制剂
免责声明:本资产仅供美国媒体专业人士使用。Sonrotoclax 是一种安全性和有效性尚未确定的试验药物。什么是 SONROTOCLAX?Sonrotoclax 是一种具有差异化特征的试验性小分子 B 细胞淋巴瘤 2 (BCL2) 抑制剂

Alpha-1蛋白酶抑制剂疗法
• Aralast NP(α-1 蛋白酶抑制剂) • Glassia(α-1 蛋白酶抑制剂) • Prolastin-C(α-1 蛋白酶抑制剂) • Zemaira(α-1 蛋白酶抑制剂) α-1 抗胰蛋白酶缺乏症 (AATD) 是一种遗传性疾病,其特征是血清和肺中 α-1 抗胰蛋白酶 (AAT) 浓度不足。这种缺乏会导致肺部丝氨酸蛋白酶(如中性粒细胞弹性蛋白酶)和 AAT 之间的不平衡。中性粒细胞弹性蛋白酶会破坏弹性蛋白,而 AAT 会防止弹性蛋白降解。这种不平衡会导致肺结缔组织破坏和早发性肺气肿的发展。AATD 还会影响肝细胞并导致肝损伤、肝硬化或肝功能衰竭。严重的 AATD 尚未得到充分认识,已知约有 100,000 名美国人患有该病。AATD 的诊断依赖于对个体血清 AAT 水平的实验室评估。 AAT 可通过放射免疫扩散法、火箭免疫电泳法或比浊法进行评估。不同的测试具有略微不同的正常范围,并且检测 AAT 缺乏症的临界点因测试而异。使用静脉注射 alpha-1 蛋白酶抑制剂的慢性增强疗法用于治疗患有先天性 AATD 和临床明显肺气肿的个体,以减缓疾病的进展。治疗的目标是通过将 AAT 水平提高到保护阈值以上来纠正中性粒细胞弹性蛋白酶的不平衡。肺部中性粒细胞弹性蛋白酶水平会因感染和香烟烟雾等刺激物而升高。影响肺功能下降的一个重要风险因素是当前吸烟。因此,仅建议以前吸烟或不吸烟的人使用增强疗法。AAT 增强疗法的安全性和有效性数据质量较差,并且报告结果没有显著差异,或者在某些情况下,肺功能下降。然而,美国胸科学会/欧洲呼吸学会(2003 年)和加拿大胸科学会(2012 年)已发布指导意见,建议对中度气流阻塞(FEV 1 为预测值的 30-65%)和肺功能快速下降(FEV 1 变化 > 120 毫升/年)的个体进行增强治疗。这些指南并不建议对无肺气肿的 AATD 个体或气道阻塞轻度或重度的个体进行增强治疗。α-1 蛋白酶抑制剂来源于混合人血浆,可能含有微量的 IgA。已知有 IgA 抗体的个体(可能存在于选择性或重度 IgA 缺乏的个体中)发生潜在严重超敏反应和过敏反应的风险更大。由于有严重超敏反应的风险,α-1 蛋白酶抑制剂禁用于有抗 IgA 抗体的个体。

ly3962673 kras g12d 抑制剂
缩写:EGFR=表皮生长因子受体;ERK=细胞外信号调节激酶;G12D=12 位甘氨酸突变为天冬氨酸;GDP=二磷酸鸟苷;GTP=三磷酸鸟苷;HRAS=Harvey 大鼠肉瘤病毒;KRAS=Kirsten 大鼠肉瘤病毒;LY=LY3962673;MEK=丝裂原活化蛋白激酶;NRAS=神经母细胞瘤 RAS 病毒致癌基因同源物;RAF=快速加速纤维肉瘤;RTK=受体酪氨酸激酶。参考文献:1. Kano Y 等人。Nat Commun. 2019;10(1):224。2. Hofmann MH 等人。Cancer Discov. 2022;12(4):924-937。3. Ostrem JML 等人。Nat Rev Drug Discov。2016;15(11):771-785。4. Gong X 等人。海报发表于:AACR 2024。摘要 3316。5. Iyer C 等人。海报发表于:AACR 2024。摘要 B115。

血管紧张素转换酶(ACE)抑制剂
案例表现是一名61岁的女性,患有哮喘,季节性过敏和高血压的过去病史,患有严重的10/10弥漫性情节性腹部疼痛。疼痛最初是在她的ED访问前2个月开始的,在进食后恶化并减轻了旋转。相关的症状包括恶心,咳嗽和偶尔呕吐。患者对系统的审查对她的呕吐或粪便,发烧,发冷,胸痛或呼吸短暂的血液不利。在介绍之前的药物包括需要根据需要,每天40毫克Atorvastatin和lisinopril每天20毫克。她已经出现了一个同样的症状,曾向她的初级保健医生和ED介绍。当时,她的代谢面板和肝功能测试正常,并且在接受IV液体后被送回家。回到ED时,她的生命体征是稳定的。体格检查是良性的,除了在所有4个象限中都有轻度嫩腹部。她的完整血液工作仅对于中性粒细胞预占68%的白细胞数量为12.20千/UL(4.31-1016千/ul)而言是显着的。腹部和骨盆的 CT扫描显示,空肠的粘膜水肿以及有关肠炎肠炎的肠系膜血管的影响。 与诊断放射科医生讨论了此病例,鉴于患者的赖诺普利使用,他提出了ACEI诱导的血管性水肿。 呼叫的外科医生同意这项评估,并决定继续进行保守管理。 她的lisinopril被切换到氢氯噻嗪。CT扫描显示,空肠的粘膜水肿以及有关肠炎肠炎的肠系膜血管的影响。与诊断放射科医生讨论了此病例,鉴于患者的赖诺普利使用,他提出了ACEI诱导的血管性水肿。呼叫的外科医生同意这项评估,并决定继续进行保守管理。她的lisinopril被切换到氢氯噻嗪。持有患者的丽二吡啶药物,并用肠休息为NPO,并用吗啡和对乙酰氨基酚对她的疼痛进行了治疗。第二天,患者的症状已经完全解决,她的腹部检查是良性的。她被从医院出院,一周后在诊所中随访,并保持症状。她的电子病历中的过敏清单已更新为ACEI。

risankizumab,一种白介素-23抑制剂,用于中等...
结构化的抽象问题:risankizumab(Skyrizi,Abbvie Pharmaceuticals,CA),旧金山,加利福尼亚州),白介素(IL)-23抑制剂?设计:进行了两个多中心,随机,双盲,安慰剂对照试验,前进和动力。预先试验包括未耐受或对≥1种经过的常规疗法(例如,皮质类固醇,5-ASA产物)或生物逻辑的患者没有足够的反应,而动机试验仅包括先前生物学失败的患者。在两项试验中,暴露于Ustekinumab(Stelara; Janssen Pharmaceuticals,Beerse,Belgium)的患者均在两项试验中均为20%。该试验是在35天的筛查期间进行的,诱导期为12周。他们需要进行回肠cosonoscopicy在筛查期间确定eligibil,然后在第12周。有临床反应的患者继续参加了