XiaoMi-AI文件搜索系统
World File Search SystemInsertions
LPA 海报 ASGCT 2023
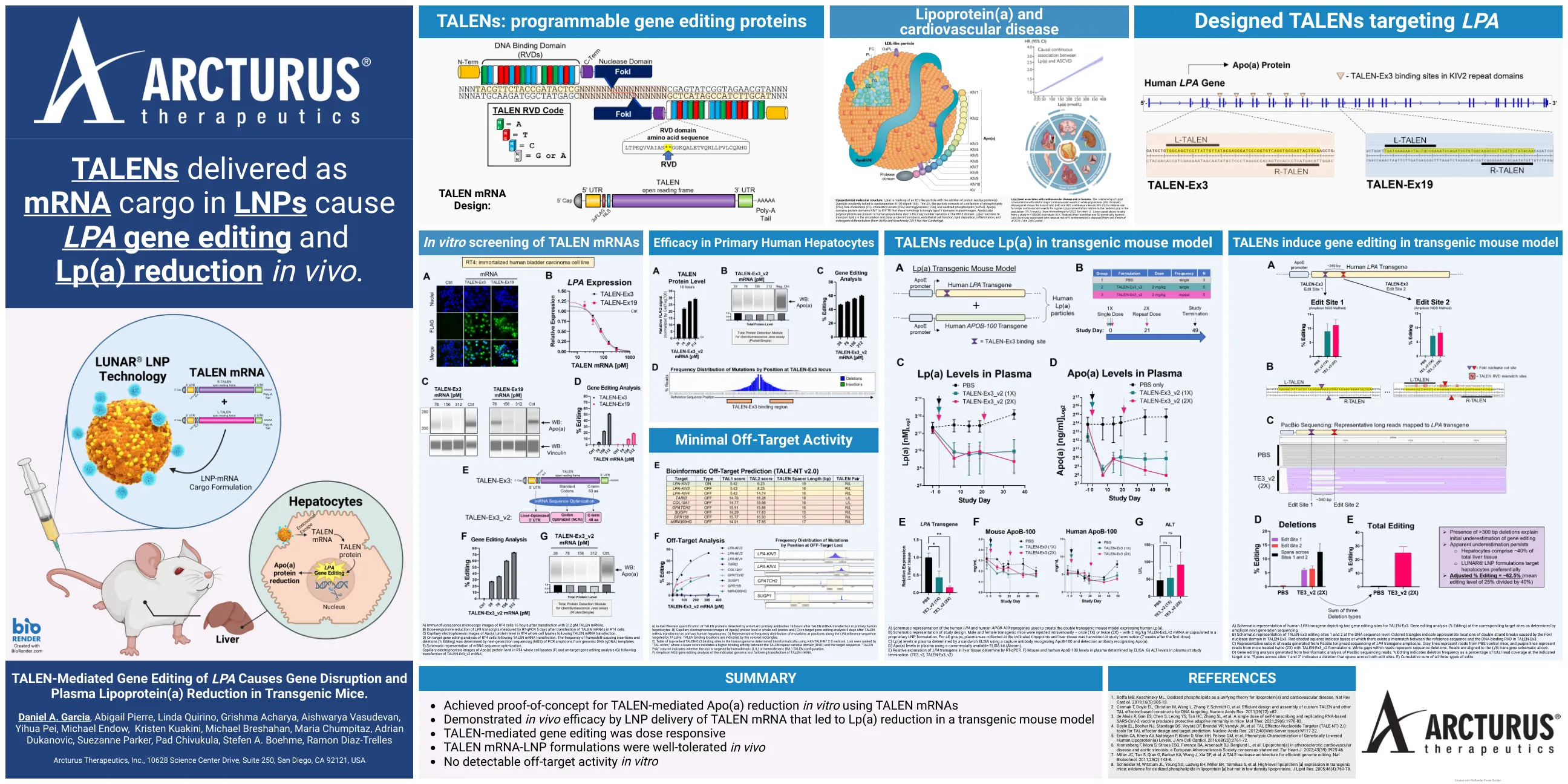
A) RT4 细胞转染 312 pM TALEN mRNA 16 小时后的免疫荧光显微镜图像。B) RT4 细胞转染 TALEN mRNA 5 天后,通过 RT-qPCR 测量 LPA 转录本的剂量反应性减少。C) TALEN mRNA 转染后 RT4 全细胞裂解物中 Apo(a) 蛋白水平的毛细管电泳图像。D) TALEN mRNA 转染后 RT4 细胞的靶向基因编辑分析。通过基因组 DNA (gDNA) 模板的 PCR 扩增子的下一代测序 (NGS) 确定导致移码的插入和缺失的频率 (编辑百分比)。E) mRNA 序列优化的示意图。转染 TALEN-Ex3_v2 mRNA 后 RT4 全细胞裂解物中 Apo(a) 蛋白水平的毛细管电泳图像 (F) 和靶向基因编辑分析 (G)。

TruSight 肿瘤学综合 - accessdata.fda.gov
– 如果肿瘤含量(按面积计算)低于 20%,则可能无法可靠地检测到体细胞驱动突变。 • 尚未评估 TSO Comprehensive 在接受器官或组织移植的患者样本中的表现。 • 尚未确定变异等位基因频率 (VAF) 低于 5% 的小 DNA 肿瘤分析变异的准确性。 • 仅在脑组织中确定了 RNA 中 EGFRvIII 剪接变异的准确性。 尚未确定其他组织类型中 EGFRvIII 的准确性。 • 已在细胞系中确定了插入 1-2 个碱基对和二核苷酸重复的检测限 (LoD) • 污染检测可能受以下因素影响: – 在高度重排的基因组中,存在缺失和杂合性缺失,TSO Comprehensive 软件可能会错误地

利用长阅读的组件和机器学习来增强短阅读转座
图1:(a)描述用于检测非参考TE 97插入的读取图信息的图表。简短读取与参考基因组对齐,并读取98,其中一个在对参考基因组中读取,而另一个读取为TE序列99(不一致的映射读取)或读取一对在参考和TE 100序列之间分配的,而TE 100序列(分裂读数)被量化。(b)Teforest管道的概述。输入和输出101个文件显示在椭圆形中,管道中的重要分支点显示在钻石中,管道的102个计算步骤显示在矩形中。(c)IGV中显示的TE插入周围的103个对齐模式的示例。将映射到基因组中其他地方的TE序列104中以颜色显示。105

DNA组装与合成生物学
用户友好的DNA工程方法可以实现多个PCR片段组件,核苷酸序列改变和定向克隆。靶DNA分子和克隆载体由PCR产生,而相邻片段之间具有6-10个同源性碱基。pCR引物包含一个二氧化神经菌残基(DU),该残基(DU)在同源性区域的3´末端,可以容纳核苷酸取代,插入和/或缺失。然后使用引物用离散的重叠片段扩增向量和靶DNA,这些片段在两端都包含DU。随后使用用户酶对PCR片段进行处理会在每个DU上产生一个单个核苷酸间隙,从而导致PCR片段侧翼,侧面有SS延伸,使定制DNA分子的无缝和方向组装成线性化的载体。多碎片组件和/或各种诱变变化。
第1页,共4页卫生技术评论
Guardant360®是一种基于定性的下一代测序测试,它使用靶向高吞吐杂交捕获技术检测74个基因中的单核苷酸变体(SNV),插入和删除(Indels),拷贝数扩增(CNAS)中的18(18)Genes in 6(18)Genes in六(6)(6)(6)(6)。Guardant360利用无细胞的DNA(CFDNA)从无细胞DNA血液收集管(BCT)中收集的外周全血的血浆中。Guardant360提供了基因组结果,包括使用常规的血液抽血在实验室的样品收据中7天内在7天内进行基因组结果,从而消除了仅依靠组织测试的需求。Guardant360为晚期固体癌症患者提供明智的治疗决策,并在一线治疗或进展前确定患者的治疗选择或临床试验。

肽融合提高主要编辑效率 1
Prime editing 是一种基于 CRISPR 的“搜索和替换”技术,可在没有双链断裂 (DSB) 或供体 DNA 模板 1 的情况下,在哺乳动物细胞中介导靶向 32 插入、删除和所有可能的碱基对碱基转换。Prime editing 34 酶 (PE2) 由与工程逆转录酶 (RT) 融合的 SpCas9 切口酶组成。35 PE2 通过 Prime editing 向导 RNA (pegRNA) 被招募到目标位点,该 RNA 除了标准基因组靶向间隔区和 SpCas9 结合发夹结构外,还包含 3' 序列,37 该序列充当融合 RT 的模板,以在一条切口 DNA 链上合成编程的 DNA 序列。当细胞 DNA 修复机制修复断裂的链时,这种 RT-39 延伸片段会与未编辑的片段竞争,而编辑后的序列有时会取代基因组中的原始序列 1,2。41


NEBNEXT微生物组DNA富集试剂盒E2612手册
用户友好的DNA工程方法可以实现多个PCR片段组件,核苷酸序列改变和定向克隆。靶DNA分子和克隆载体由PCR产生,而相邻片段之间具有6-10个同源性碱基。pCR引物包含一个二氧化神经菌残基(DU),该残基(DU)在同源性区域的3´末端,可以容纳核苷酸取代,插入和/或缺失。然后使用引物用离散的重叠片段扩增向量和靶DNA,这些片段在两端都包含DU。随后使用用户酶对PCR片段进行处理会在每个DU上产生一个单个核苷酸间隙,从而导致PCR片段侧翼,侧面有SS延伸,使定制DNA分子的无缝和方向组装成线性化的载体。多碎片组件和/或各种诱变变化。

DNA组装与合成生物学
用户友好的DNA工程方法可以实现多个PCR片段组件,核苷酸序列改变和定向克隆。靶DNA分子和克隆载体由PCR产生,而相邻片段之间具有6-10个同源性碱基。pCR引物包含一个二氧化神经菌残基(DU),该残基(DU)在同源性区域的3´末端,可以容纳核苷酸取代,插入和/或缺失。然后使用引物用离散的重叠片段扩增向量和靶DNA,这些片段在两端都包含DU。随后使用用户酶对PCR片段进行处理会在每个DU上产生一个单个核苷酸间隙,从而导致PCR片段侧翼,侧面有SS延伸,使定制DNA分子的无缝和方向组装成线性化的载体。多碎片组件和/或各种诱变变化。

利用 DNA-PKcs 抑制技术在人类原代细胞中通过同源性定向修复实现高效转基因整合
基于核酸酶的基因组编辑的治疗应用将受益于通过同源定向修复 (HDR) 进行转基因整合的改进方法。为了提高 HDR 效率,我们筛选了六种 DNA 依赖性蛋白激酶催化亚基 (DNA-PKcs) 的小分子抑制剂,DNA-PKcs 是替代修复途径非同源末端连接 (NHEJ) 中的关键蛋白,可产生基因组插入/缺失 (INDEL)。从这次筛选中,我们确定 AZD7648 是最有效的化合物。使用 AZD7648 可显著增加 HDR(高达 50 倍)并同时降低各种治疗相关的原代人类细胞类型中不同基因组位点的 INDEL。在所有情况下,HDR 与 INDEL 的比率均显著增加,并且在某些情况下,实现了无 INDEL 的高频 (>50%) 靶向整合。这种方法有可能提高基于细胞的疗法的治疗效果并扩大靶向整合作为研究工具的使用范围。