XiaoMi-AI文件搜索系统
World File Search SystemK562
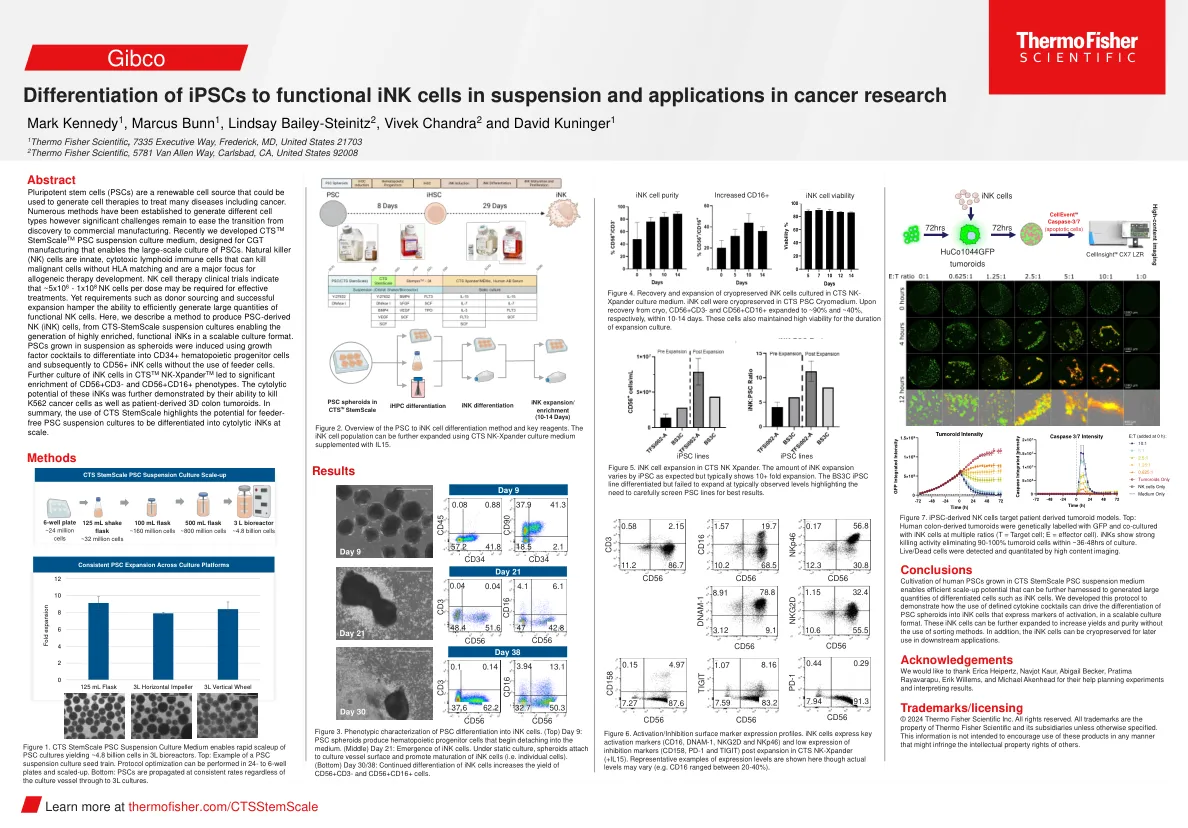
差异化-IPSCS功能 - 互联 - 悬崖 - casser ...
stemscale TM PSC悬浮培养基,设计用于CGT制造,可实现PSC的大规模培养。天然杀伤(NK)细胞是先天的,细胞毒性的淋巴免疫细胞,可以杀死恶性细胞而无需HLA匹配,并且是同种异体治疗发展的主要重点。nk细胞疗法临床试验表明,有效治疗可能需要〜5x10 6-1x10 8 nk细胞。但诸如捐助者采购和成功扩展之类的要求妨碍了有效产生大量功能性NK细胞的能力。在这里,我们描述了一种产生PSC衍生的NK(墨水)细胞的方法,该方法从CTS-STAMScale悬浮培养物中,使能够以可扩展的培养格式产生高度富集的功能性油墨。PSC在悬浮液中生长,因为使用生长因子鸡尾酒诱导球体,以区分CD34+造血祖细胞,然后在不使用进料细胞的情况下转化为CD56+墨水细胞。CTS TM NK-Xpander TM中墨水细胞的进一步培养导致CD56+CD3-和CD56+CD16+表型的显着富集。这些墨水的细胞溶性通过它们杀死K562癌细胞以及患者来源的3D结肠肿瘤的能力进一步证明了这些墨水。总而言之,CTS茎扫描的使用突出了馈线 - 游离PSC悬浮培养物的潜力,以分为大规模的细胞溶解油墨。

TYRO3 敲低可抑制髓系白血病细胞的生长
摘要。背景/目的:TYRO3 是受体酪氨酸激酶 TAM 家族 (TYRO3、AXL 和 MERTK) 的成员。虽然已报道了激活的 AXL 和 MERTK 在白血病细胞生长中的作用,但 TYRO3 的影响尚未确定。因此,我们研究了 TYRO3 敲低对白血病细胞系生长的影响。材料和方法:本研究使用了三种表达 TYRO3 蛋白的人类白血病细胞系 (纯红细胞白血病衍生的 AA、OCI/AML2 和 K562)。为了诱导 TYRO3 敲低,使用电穿孔系统转染针对 TYRO3 的小干扰 RNA (siRNA)。通过比色测定评估细胞生长。通过免疫印迹检查各种信号蛋白的表达水平和激活。通过微阵列分析检查 TYRO3 敲低后综合基因表达的变化。结果:TYRO3 敲低抑制了所测试的白血病细胞系中的细胞生长。此外,敲低还抑制了 AA 细胞中的信号转导和转录激活因子 3 的磷酸化,以及 AA 和 OCI/AML2 细胞中的细胞外信号调节激酶 (ERK) 1/2;两者都是 TYRO3 信号传导的下游分子。TYRO3 敲低还抑制了所有细胞系中 survivin 的表达。TYRO3 敲低强烈抑制了 TYRO3 mRNA 表达,但没有抑制 AXL 和 MERTK 的表达。此外,TYRO3 敲低抑制了 ERK 下游分子细胞周期蛋白 D1 mRNA 的表达。结论:TYRO3 在白血病细胞生长中发挥作用,是白血病的潜在治疗靶点。

对改变转录因子间距的自然插入和缺失的系统分析可识别耐受性和敏感的转录
基因表达的抽象调节需要在启动子和增强子上对序列特异性转录因子(TFS)的联合结合。先前的研究表明,TF结合位点之间间距的改变会影响启动子和增强子活性。然而,由于自然发生的插入和删除(Indels)导致的TF间距改变的重要性尚未系统地分析。为了解决这个问题,我们首先表征了通过ChIP-Seq(Chro-Matin免疫沉淀测序)确定的人类K562细胞中73 TF的全基因组间距关系。我们发现了协作因素之间放松的间距的主要模式,其中包括45个TFS专门与其结合伴侣展示了放松的间距。接下来,我们利用了遗传多样的小鼠菌株和人个体提供的数百万个indels来研究间距改变对TF结合和局部组蛋白乙酰化的影响。这些分析表明,与直接影响TF结合位点的遗传变异相比,通常可以容忍自然存在的插入的间距改变。为了实验验证这一预测,我们在巨噬细胞系中的六个内源基因组基因座上引入了PU.1和C/EBPβ结合位点之间的合成间距改变。在这些位置,PU.1和C/EBPβ的合作结合明显,可耐受的间距的变化范围从5 bp增加到> 30 bp的降低。总的来说,这些发现对理解增强子选择的机制以及对非编码遗传变异的解释具有影响。

预测基于 CRISPR-Cas9 的表观基因组编辑的效果
摘要表观遗传调控协调哺乳动物转录,但它们之间的功能联系仍然难以捉摸。为了解决这个问题,我们使用来自 13 种 ENCODE 细胞类型的表观基因组和转录组数据来训练机器学习模型,以预测组蛋白翻译后修饰 (PTM) 的基因表达,对于大多数细胞类型,实现了 ∼0.70 −0.79 的转录组范围相关性。我们的模型重现了组蛋白 PTM 和表达模式之间的已知关联,包括预测转录起始位点 (TSS) 附近的组蛋白亚基 H3 赖氨酸残基 27 (H3K27ac) 的乙酰化会显著提高表达水平。为了通过实验验证这一预测,并研究 H3K27ac 的天然沉积与人工沉积对表达的影响,我们将合成的 dCas9-p300 组蛋白乙酰转移酶系统应用于 HEK293T 细胞系中的 8 个基因和 K562 细胞系中的 5 个基因。此外,为了便于建立模型,我们执行 MNase-seq 来绘制 HEK293T 中全基因组核小体占有水平。我们观察到,我们的模型在准确排序基因对 dCas9-p300 系统的相对倍数变化方面表现良好;然而,与根据其天然表观遗传特征预测跨细胞类型的表达相比,它们对单个基因内倍数变化进行排序的能力明显减弱。我们的研究结果强调,我们需要更全面的基因组规模表观基因组编辑数据集,更好地理解表观基因组编辑工具所做的实际修改,以及改进因果模型,以便更好地从内源性细胞测量转移到扰动实验。这些改进将共同促进理解和可预测地控制动态人类表观基因组的能力,以及对人类健康的影响。

含T-2毒素的pH敏感脂质体的制备……
摘要:T-2毒素为A型单端孢霉烯族毒素。为了降低T-2毒素的副作用并提高其肿瘤靶向性,本研究制备并表征了T-2毒素pH敏感脂质体(LP-pHS-T2)。以T-2毒素为对照,采用3-(4,5-二甲基噻唑-2)-2,5-二苯基四唑溴化物法检测LP-pHS-T2对A549、Hep-G2、MKN-45、K562和L929细胞系的细胞毒性。研究了LP-pHS-T2对Hep-G2细胞的凋亡和迁移影响。LP-pHS-T2的制备工艺涉及以下参数:二棕榈酰磷脂酰胆碱:二油酰磷脂酰乙醇胺,1:2;总磷脂浓度20 mg/ml,磷脂:胆固醇3:1,4-(2-羟乙基)-1-哌嗪乙磺酸缓冲液(pH 7.4),10 ml,药脂比2:1,超声10 min后挤压,包封率达95±2.43%。挤压后LP-pHS-T2平均粒径为100 nm,透射电镜观察显示LP-pHS-T2呈圆形或椭圆形,大小均匀。释放曲线呈现两阶段下降趋势,前6 h T-2毒素快速渗漏(释放量~20%),随后持续释放至48 h(释放量~46%),48-72 h渗漏率增加(释放量~76%),72 h时达到最低。当LP‑pHS‑T2浸泡在0.2 mol/l磷酸二钠‑磷酸二氢钠缓冲液(pH 6.5)中时,释放速度明显加快,释放率可达91.2%,表现出较强的pH敏感性。抗肿瘤试验表明,LP‑pHS‑T2能够促进Hep‑G2细胞凋亡,抑制其迁移。本研究为基于T‑2毒素的抗癌药物的开发提供了一种新方法。
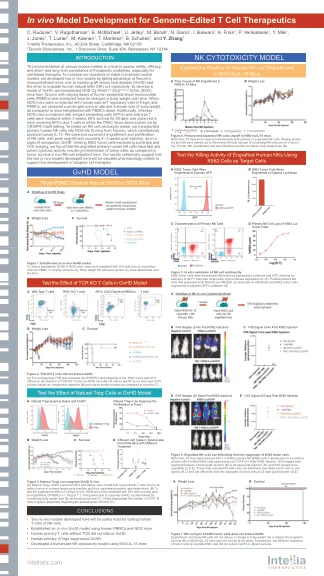
基因组编辑 T 细胞治疗的体内模型开发
实施各种小鼠模型对于评估治疗方式的安全性、有效性以及短期和长期持久性至关重要,尤其是对于基于细胞的疗法。为了增加我们可行的人源化小鼠模型库,我们利用 Taconic 的免疫缺陷小鼠开发了两种体内模型:一种用于监测移植物抗宿主病 (GvHD),另一种用于评估人类自然杀伤 (NK) 细胞的细胞毒性。为了开发 GvHD 模型,我们向 Taconic 的 NOD.Cg- Prkdc scid Il2rg tm1Sug / JicTac (NOG) 小鼠移植了不同剂量的人类外周血单核细胞 (PBMC),并监测小鼠体重随时间的变化。当 NOG 小鼠同时注射人类天然 T 调节细胞 (nTregs) 和 PBMC 时,与仅移植 PBMC 的小鼠相比,我们观察到存活率延长且体重减轻较慢。此外,同时注射抗原呈递细胞 (APC) 和野生型 T 细胞的 NOG 小鼠在 3 周内就奄奄一息,而接受 APC 加 T 细胞(其中 TRAC 基因座通过 CRISPR / Cas9 编辑被敲除)的小鼠在 90 天内存活率达到 80%。为了建立 NK 细胞毒性模型,我们将原代人类 NK 细胞移植到 Taconic 的 NOG-hIL15 小鼠中,该小鼠组成性产生人类 IL-15。我们观察到 NK 细胞的成功植入和增殖,峰值植入发生在注射后 4 - 5 周,没有异种 GvHD 的迹象。利用表达荧光素酶的 K562 肿瘤细胞和 IVIS 成像,我们发现植入的原代人类 NK 细胞具有快速而强大的细胞毒活性,与非 NK 细胞植入小鼠的肿瘤存活率相比,可消除肿瘤细胞。我们的研究结果共同表明,这里开发的两种体内模型将成为支持过继细胞疗法发展的有价值的药理学模型。

HLA-A/B/C敲除电穿孔套件
说明HLA-A/B/C敲除电穿孔套件适用于通过电穿孔的细胞系和原代T细胞工程。该套件既包含Cas9酶(链球菌)和靶向HLA-A/B/C(人白细胞抗原)的GRNA。该套件足以设计高达500万个原代T细胞。背景HLA(人白细胞抗原)-a,b和c是MHC的三种主要类型(主要的组织相容性复合物)1类跨膜蛋白。它们与β2微球蛋白蛋白(由B2M基因编码)形成异二聚体。MHC 1类分子表现出短多肽,通常在长7-11个氨基酸之间,以识别为“自我”或“非自身”的免疫系统。HLA-C存在于所有细胞中,并且由于HLA-C基因的多样性而作为几种单倍型存在。c*08:02代表一种这样的单倍型。HLA I类将新抗原衍生的肽呈现到细胞表面,从而通过TCR(T细胞受体)识别出T细胞的识别。 癌症免疫疗法一直在使用该机制,方法是表达能够识别特定癌症免疫原子的TCR。 在2016年,HLA-C*08:02限制性TIL(肿瘤浸润淋巴细胞)在肺癌中靶向KRAS(Kirsten大鼠肉瘤病毒)G12D突变,导致阳性结果。 在转移性胰腺癌患者中采用了类似的方法,并导致该疾病的消退。 HLA-C*08:02限制性TIL对其他新抗原的TCR的研究可能对癌症治疗有益。 应用程序HLA I类将新抗原衍生的肽呈现到细胞表面,从而通过TCR(T细胞受体)识别出T细胞的识别。癌症免疫疗法一直在使用该机制,方法是表达能够识别特定癌症免疫原子的TCR。在2016年,HLA-C*08:02限制性TIL(肿瘤浸润淋巴细胞)在肺癌中靶向KRAS(Kirsten大鼠肉瘤病毒)G12D突变,导致阳性结果。在转移性胰腺癌患者中采用了类似的方法,并导致该疾病的消退。HLA-C*08:02限制性TIL对其他新抗原的TCR的研究可能对癌症治疗有益。应用程序K562细胞是HLA I和II类负的,使其成为引入和研究特定单倍型响应的理想细胞模型。hla在供体细胞和个体之间的不匹配可以导致免疫排斥反应,一种选择是敲除内源性HLA,从而使细胞被更广泛地普遍使用。

Sanjay Ghosh
使用酵母作为模型系统来表征硝化应激反应,增加了证据的数量,这表明反应性氮物种(RNSS)和一氧化氮(NO)本身会影响细胞的氧化还原状态,例如氧化应激和修饰细胞蛋白,可逆地或不可逆地修饰细胞蛋白。酵母是研究细胞中反应性氮种的作用的出色模型系统。目前,我们正在研究BZIP转录因子ATF1和PCR1在硝化应激中的作用。研究亚硝化应激对酿酒酵母的线粒体呼吸链超复合物的影响表征NO和RNS对细胞死亡机制的影响NO和RNS对慢性骨髓骨髓性K562细胞系和MCF7细胞中的NO和RN对细胞死亡机制的作用。分泌植物学真菌巨摩托菌的分类分析在固态培养中生长。,我们开发了一种生物处理方法,用于使用巨型球虫中的固体发酵生产内糖酸酶和木烷酶。研究弧菌霍乱中的硝化应力反应机制。9。研究指南:注册博士学位主管,部门加尔各答大学生物化学,2001年3月。授予博士学位的研究人员人数学位:追求M.Phil./博士学位的十八(18)个研究人员人数:第四(4)届:1)Chirandeep Dey,B.Sc。&M.Sc.在动物学中,UGC-NET SRF 2)Ayantika Sengupta,学士学位&M.Sc.在动物学中,CSIR-NET SRF 3)SANCHITA BISWAS,B.SC。动物学和硕士在生物化学中,CSIR-NET SRF 4)SHUDDHASATTWA SAMADDAR,B.SC。微生物学和硕士学位 在生物化学中,DBT-SRF 5)Sourav Mukherjee,硕士 生物技术,项目实习生前博士学生:1)Rajib Sengupta博士,学士学位 化学硕士 生物化学博士学位在2007年颁发的奖学博士研究生授予,在匹兹堡大学外科Detcho A. Stoyanovsky教授的监督下,匹兹堡大学后研究员,在Karolinska Institutet的Karolinska Institutet的Biiochemist和Biophysics教授Arne Holmgren教授的监督下微生物学和硕士学位在生物化学中,DBT-SRF 5)Sourav Mukherjee,硕士生物技术,项目实习生前博士学生:1)Rajib Sengupta博士,学士学位 化学硕士 生物化学博士学位在2007年颁发的奖学博士研究生授予,在匹兹堡大学外科Detcho A. Stoyanovsky教授的监督下,匹兹堡大学后研究员,在Karolinska Institutet的Karolinska Institutet的Biiochemist和Biophysics教授Arne Holmgren教授的监督下生物技术,项目实习生前博士学生:1)Rajib Sengupta博士,学士学位化学硕士 生物化学博士学位在2007年颁发的奖学博士研究生授予,在匹兹堡大学外科Detcho A. Stoyanovsky教授的监督下,匹兹堡大学后研究员,在Karolinska Institutet的Karolinska Institutet的Biiochemist和Biophysics教授Arne Holmgren教授的监督下化学硕士生物化学博士学位在2007年颁发的奖学博士研究生授予,在匹兹堡大学外科Detcho A. Stoyanovsky教授的监督下,匹兹堡大学后研究员,在Karolinska Institutet的Karolinska Institutet的Biiochemist和Biophysics教授Arne Holmgren教授的监督下

基于细胞的抗慢病毒转导人工 APC,可从各种细胞来源有效生成 CAR-T 细胞
摘要 背景 嵌合抗原受体 T 细胞 (CAR-T) 的过继细胞疗法已经成为某些侵袭性 B 细胞恶性肿瘤患者的标准治疗方法,并有望在未来改善许多其他癌症患者的护理。然而,CAR-T 细胞疗法的高制造成本对其更广泛的临床应用构成了重大障碍。CAR-T 生产的主要成本驱动因素包括用于 T 细胞活化的一次性试剂和临床级病毒载体。起始材料中存在不同数量的污染单核细胞对 CAR-T 制造构成了额外的挑战,因为它们会阻碍 T 细胞刺激和转导,导致制造失败。方法我们创建了基于 K562 的人工抗原呈递细胞 (aAPC),具有基因编码的 T 细胞刺激和共刺激,这是 T 细胞活化的取之不尽的来源。我们还使用 CRISPR-Cas9 基因编辑核酸酶破坏了这些 aAPC(aAPC- Δ LDLR)上低密度脂蛋白受体 (LDLR) 的内源性表达,以防止意外慢病毒转导并避免转导过程中对病毒载体的吸收效应。使用各种 T 细胞来源,我们通过基于 aAPC- Δ LDLR 的激活产生了 CD19 导向的 CAR-T 细胞,并在体外和体内测试了它们对 B 细胞恶性肿瘤的抗肿瘤效力。结果我们发现 aAPC- Δ LDLR 上缺乏 LDLR 表达会导致 CAR-T 生产过程中对慢病毒转导产生抗性。使用 aAPC- Δ LDLR,我们甚至可以从未纯化的起始材料(如外周血单核细胞或未经处理的白细胞分离术产品,其中含有大量单核细胞)中实现 CAR-T 细胞的有效扩增。我们通过基于 aAPC- Δ LDLR 的扩增产生的 CD19 定向 CAR-T 细胞在急性淋巴细胞白血病和 B 细胞淋巴瘤的临床前模型中表现出强大的抗肿瘤反应。结论我们的 aAPC- Δ LDLR 代表了一种用于制造慢病毒转导 T 细胞的有吸引力的方法

MicroRNA-1246 通过靶向 AXIN2 和 GSK-3β...
背景与目的:化疗在白血病治疗中起着重要作用。化疗引起的多药耐药性 (MDR) 往往导致治疗失败和疾病复发。微小 RNA (miRNA) 已被证实是致癌作用的关键组成部分,包括肿瘤细胞的化学耐药性,但这一点尚未完全了解。在本研究中,我们旨在确定潜在的候选 miRNA miR-1246,并揭示其在白血病细胞化学耐药中的调控作用。方法:通过微阵列分析选择候选 miRNA,通过生物信息学工具筛选并通过逆转录定量聚合酶链反应 (RT-qPCR) 进行验证。检测转染 miR-1246 类似物或抑制剂后白血病细胞的化疗耐药表型,包括细胞存活率、凋亡、阿霉素 (ADM) 外排和体内致癌性,并检测是否接受 ADM 处理,以明确 miR-1246 与化疗耐药之间的关系。通过 RT-qPCR、Western blot 和双荧光素酶报告基因检测,检测相关基因的表达,探讨 miR-1246 在化疗耐药中的潜在调控机制。结果:miR-1246 在化疗耐药的白血病 K562/ADM 细胞、HL-60/RS 细胞和复发性原发性白血病细胞中的表达显著增高。 miR-1246的缺失抑制了化疗耐药白血病细胞的增殖、诱导了细胞凋亡、改变了细胞周期分布、抑制了ADM的流出,而miR-1246的过表达在化疗敏感白血病细胞中则表现出相反的作用。生物信息学预测和荧光素酶检测均表明AXIN2和糖原合酶激酶3β(GSK-3β)是白血病细胞中miR-1246的直接作用靶点。抑制miR-1246可以上调AXIN2和GSK-3β并使Wnt /β-catenin通路失活,同时抑制β-catenin的表达,并进一步影响化疗耐药白血病细胞中P糖蛋白(P-gp)的表达。结论: miR-1246 的缺失通过负向调控 AXIN2 和 GSK-3 β,使 Wnt/β-catenin 通路失活并抑制 P-gp 表达,从而减弱了 MDR 白血病细胞的化疗耐药能力,这意味着靶向 miR-1246-AXIN2/GSK-3β-Wnt/β-catenin 轴可能有利于克服复发和难治性白血病患者的化疗耐药性。