XiaoMi-AI文件搜索系统
World File Search SystemKauffman

路易斯·考夫曼传记
考夫曼的研究领域是代数拓扑,尤其是低维拓扑和结理论,以及它们与数学物理和自然科学的关系。20 世纪 70 年代早期,他对高维结和高维流形上的奇异结构的研究使用了分支覆盖构造的概括,对于通过 Brieskorn 簇和代数奇点链表达的这些结构的拓扑理解至关重要。这些非标准可微结构的构造至今仍是个谜,并且肯定与基础物理学有关——就像 Brieskorn 研究的流形一样。考夫曼于 1980 年发现了亚历山大-康威多项式的状态求和模型,并于 1985 年发现了琼斯多项式的括号多项式状态模型。这些状态模型构成了分区函数在结不变量构造中的首次直接应用。在括号多项式模型中,考夫曼表明,这种状态总和是统计力学中 Potts 模型的一个版本 - 转换为结点图。他发现了原始琼斯多项式的二变量泛化,称为半定向或考夫曼多项式。自从这些发现以来,他的工作主要针对结点和链接的新不变量的结构。括号模型使考夫曼、Murasugi 和(独立)Thistlethwaite 证明了 Tait 猜想,即减少交替链接投影的交叉数的拓扑不变性。他在虚拟结点理论方面的研究开辟了结点理论的新领域,并发现了许多结点和链接的新不变量。特别是,考夫曼括号中的状态结构被米哈伊尔·霍瓦诺夫 (Mikhail Khovanov) 用于创建结点的霍瓦诺夫同源理论,产生了新的和微妙的不变量。 Dye、Kauffman 和 Kaestner 利用 Manturov 的构造将 Khovanov 同源性推广到虚拟结点理论,并以此方式完成了 Rasmussen 不变量的新版本。这导致了正虚拟结点的 4 球属的确定,而 Kauffman 应用此结果获得了

Louis H. Kauffman 的出版物
Louis H. Kauffman 的出版物 1. 论文 当外壳具有可变折射率时,两个同心球体的电磁波散射。(与 M. Kerker 和 W. Farone 合作),美国光学学会杂志。56(1966 年),1053-1056。 循环分支覆盖和 $0(n)$-流形。第二届紧变换群会议论文集(马萨诸塞大学,阿默斯特,马萨诸塞州,1971 年),第一部分,第 416--429 页。 数学讲义,第 298 卷,Springer,柏林,1972 年。 链接一致性的不变量。弗吉尼亚理工学院和州立大学拓扑学会议论文集,由 Raymond R. Dickman Jr. 和 Peter Fletcher 编辑,数学讲义,第 298 卷375,Springer Verlag,柏林,1973,第 153-157 页。链接流形和周期性。美国数学会刊 79(1973),570-573。链接流形。密歇根数学杂志 21(1974),33-44。分支覆盖、开卷和结点周期性。拓扑学 13(1974),143-160。结点的乘积。美国数学会刊 80(1974),1104-1107。链接一致性的不变量。拓扑学会议(弗吉尼亚理工学院和州立大学,弗吉尼亚州布莱克斯堡,1973),第 153-157 页。数学讲义,第 10 卷375,Springer,柏林,1974。分支循环覆盖的周期性。(与 Alan Durfee 合作)数学年鉴 218(1975),第 2 期,157-174。链接的签名。(与 L. Taylor 合作)Trans. Amer. Math. Soc. 216(1976),351-365。环面和环面结的微分几何。(与 Steve Jordan 合作)Delta (Waukesha) 6(1976),第 1 期,1-15。一个中心示例研讨会。(与 Steve Jordan 合作)Internat. J. Math. Ed. Sci. Tech. 7(1976),351-365。浸入和模 2 二次型。(与 Tom Banchoff 合作)Amer. Math. Monthly 84

Kauffman模型中关闭与开放式演变之间因果关系的证据
进化是一个以新颖性产生为标志的高度复杂的过程,这需要个人的历史和集体组织。在本文中,我们研究了生物组织与开放式进化(OEE)之间的关系,特别关注两者之间的因果关系。为了在化学系统中提供这种因果关系的定量证据,我们应用集装理论来评估自动催化集的出现如何影响Kauffman模型中的复杂性动态。在本文的第二部分中,我们通过分析最简单的自动催化设置对Kauffman模型中复杂性动力学的影响,特别是在没有参数相关性的情况下,加强了这种猜想。通过将自动催化集解释为化学系统中的组织结构,我们的发现为研究生物组织与OEE之间的因果关系提供了第一个数值支持。这项工作代表了对OEE与生物组织之间动态关系的初步研究的一个有希望的领域,并可能会促进其在理论生物学中的联系。

建立知识以编织沿海蓝色的保护和恢复
1 Pendleton,L.,Donato,D.C.,Murray,B.C.,Crooks,S.,Jenkins,W.A.,Sifleet,S.,Craft,C.,Fourqurean,J.W.,Kauffman,J.B.估计植被沿海生态系统转化和退化的全球“蓝碳”排放。PLOS ONE,7(9),P.E43542。2 Donato,D.C.,Kauffman,J.B.,Murdiyarso,D.,Kurnianto,K.,Stidham,M。和Kanninen,M.,2011年。红树林在热带地区最丰富的森林中。自然地球科学4:293–2978。3 28国家的NDC在缓解方面包括沿海湿地的参考,59个国家包括沿海生态系统和沿海地区的适应策略(Herr,D。和Landis,E。2016)。

2025年2月12日Rory M. McDonald ...
2022决赛入围者,TUM研究卓越创新,企业家精神,领导力2019年管理学院蒂姆·德(Tim Division tim Division tim Division tim Division'' Kauffman Foundation授予Alicia Desantola($ 40,000)2014 Kauffman基金会初级教师企业家研究奖学金(35,000美元)2014年决赛入围者,在商业政策/管理学院的杰出论文,2012年Trammell/CBA基金会教学学院教学奖


Dan Ma Office电话:电子邮件:madan@smu.edu.sg(+65 ... nah fui hoon“ fiona” OUH ENG LIEH电子邮件:elouh@smu.edu.sg 80 Stamford Road ... Ma Yunshan-计算与信息系统学院 Robert Deng 研究声明
在软件服务市场中的成本效率策略:Kauffman,Robert J。的建模结果和相关实施问题;玛,丹。(2013.0)。网格,云,系统和服务的经济学:第10届国际会议,2013年Gecon,西班牙Zaragoza,2013年9月18日至20日。程序,(pp。16-26)西班牙Zaragoza:Springer Verlag International Publishing。http://dx.doi.org/10.1007/978-3-319-02414-1_2(发布)http://dx.doi.org/10.1007/978-3-319-02414-1_2(发布)

估计... -JSSM
红树林是在全球热带和亚热带沿海地区的高含量或咸淡海洋环境的潮汐区中发现的重要生态系统(Romanach等,2018)。这些栖息地支持多样化的红树林物种,例如根茎,布鲁吉埃拉,索纳蒂亚,塞里奥普斯,阿维奇尼亚和木果(Hidayah et al。,2022)。红树林在储存碳延长的持续时间方面非常出色,并且被认为是世界上最高的碳密度之一(Adame等,2020)。最近的研究(Kauffman等,2018; Taillardat,2018)强调,红树林生态系统可以储存三到五倍的碳,而碳是陆地森林的三到五倍,其土壤碳池中存储了重要部分(Zakaria&Sharma,2020)。他们也被公认为是捕获大气碳储备并减轻全球温度持续升高的最有效方法之一(Amir,2018)。尽管覆盖少于1%
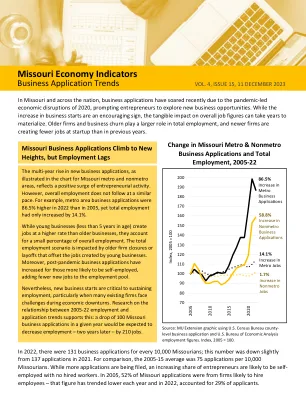
密苏里经济指标 - 业务应用趋势
在没有员工与美国劳工统计局和考夫曼基金会的调查结果保持一致的业务初创企业比例不断扩大的国家趋势中,这表明,新的企业的开业时间比往年少。例如,在2005年,新创建的密苏里州业务平均有4.7个工作。在2022年,它不到2.5个工作。这种趋势有几个原因,其中一些是由于大流行而加速的。为老年人或残疾人的个人服务,通常是提供家庭护理的单人业务,随着人口衰老的增长,这是一个越来越重要的行业。在2021 - 22年,该行业分别占密苏里州地铁和非Metro地区的新薪资业务的33%和35%。

自愿性国家含量标准
编写委员会从 2008 年到 2010 年初召开了几次会议。2009 年 10 月,一个由 Siegfried、MacDonald 和 Vredeveld 组成的小组,加上 Stephen Buckles(范德堡大学)和 Bonnie Meszaros(特拉华大学),花了三天时间密集更新和修订与 20 项标准相关的基准。在 2009 年底分发标准草案以征求公众意见后,编写委员会在 2010 年初签署了 20 项标准中 19 项的最终版本。为了回应公众对标准 14(企业家精神)的意见,编写委员会寻求更多专业知识来修订标准声明。Robert Strom(尤因马里昂考夫曼基金会)在 James O'Neill(特拉华大学)的帮助下,为这项标准提供了修订语言,委员会仅做了微小改动就接受了它。