XiaoMi-AI文件搜索系统
World File Search SystemKeap1

KEAP1 通过抑制 NSCLC 中的 PD-L1 表达来促进抗肿瘤免疫
记录版本:该预印本的一个版本于 2024 年 2 月 27 日在《细胞死亡与疾病》上发表。已发表的版本请参阅 https://doi.org/10.1038/s41419-024-06563-3 。

CRISPR/Cas9 诱导的 Keap1 缺失增强了抗...
干细胞因其再生能力而成为许多疾病治疗的有力工具,并在再生医学领域迅速推进发展,如在脑创伤方面的治疗。然而,病灶区域高氧化微环境导致99%以上的细胞死亡。本研究利用基因方法编辑间充质干细胞中的Keap1基因,并观察其抗氧化能力。首先,我们利用CRISPR/Cas9分别打乱脂肪间充质干细胞(Ad-MSCs)中Keap1的起始密码子和第376个氨基酸密码子,使Nrf2从Keap1的结合中释放出来,结果Nrf2被激活并定位到细胞核内,调控细胞的抗氧化作用。我们观察到缺少Keap1 ATG密码子的细胞表现出明显的Nrf2核定位。 H 2 O 2 处理后,除了检测到 Bax-1 表达降低和丙二醛 (MDA) 含量降低外,我们还发现 Keap1 ATG 密码子敲除细胞中 Bcl-2 表达增加,而仅在 Keap1 376 位密码子编辑细胞中观察到 PCNA 表达增加,其 Bax-1 表达低于对照细胞。我们的研究表明 Keap1 的缺失导致 Ad-MSCs 的抗氧化能力,这表明我们的策略有望提高移植后间充质干细胞的活力。本研究也是 CRISPR/Cas9 在 Ad-MSCs 中应用的前沿探索。

CTLA4 阻断可消除 KEAP1/STK11 相关的对 PD-(L)1 抑制剂的耐药性
对于晚期非小细胞肺癌 (NSCLC) 患者,与单独使用 PD-(L)1 抑制剂治疗相比,使用 CTLA4 抑制剂和 PD-1 或 PD-L1 抑制剂(下称 PD-(L)1 抑制剂)的双重免疫检查点阻断 (ICB) 具有更高的抗肿瘤活性和免疫相关毒性。然而,目前还没有经过验证的生物标志物来确定哪些患者会受益于双重 ICB 1,2 。我们在这里表明,在随机 III 期 POSEIDON 试验中,当将 PD-L1 抑制剂 durvalumab 和 CTLA4 抑制剂 tremelimumab 的双重 ICB 添加到化疗中时,患有 STK11 和/或 KEAP1 肿瘤抑制基因突变的 NSCLC 患者可从临床受益,但不能从单独使用 durvalumab 中受益 3 。公正的基因筛选发现,这两种肿瘤抑制基因的缺失是导致 PD-(L)1 抑制耐药的独立驱动因素,并表明 Keap1 的缺失是双重 ICB 疗效的最强基因组预测因子——这一发现在几种 Kras 驱动的 NSCLC 小鼠模型中得到证实。在小鼠模型和患者中,KEAP1 和 STK11 的改变都与不良的肿瘤微环境有关,其特征是抑制性髓系细胞占优势、CD8 + 细胞毒性 T 细胞耗竭,但 CD4 + 效应亚群相对保留。双重 ICB 强效作用于 CD4 + 效应细胞,并将肿瘤髓系细胞区室重新编程为表达诱导型一氧化氮合酶 (iNOS) 的杀瘤表型,其与 CD4 + 和 CD8 + T 细胞一起发挥抗肿瘤功效。这些数据支持使用双重 ICB 化学免疫疗法来减轻患有 STK11 和/或 KEAP1 改变的 NSCLC 患者对 PD-(L)1 抑制的耐药性。

PRMT5 通过甲基化 KEAP1 和抑制铁死亡降低三阴性乳腺癌的免疫治疗效果
摘要 背景 作为一种新兴的三阴性乳腺癌 (TNBC) 治疗策略,免疫治疗部分通过诱导铁死亡起作用。最近的研究表明,蛋白质精氨酸甲基转移酶 5 (PRMT5) 通过调节肿瘤微环境在多种癌症的免疫治疗中发挥不同作用。然而,PRMT5 在铁死亡过程中的作用,特别是在 TNBC 免疫治疗中的作用尚不清楚。方法 通过 IHC (免疫组织化学) 染色测量 TNBC 中的 PRMT5 表达。为了探索 PRMT5 在铁死亡诱导剂和免疫治疗中的作用,进行了功能实验。使用一组生化分析来发现潜在的机制。结果 PRMT5 促进 TNBC 中的铁死亡抗性,但削弱非 TNBC 中的铁死亡抗性。从机制上看,PRMT5选择性甲基化KEAP1,从而下调NRF2及其下游靶标,这些靶标可分为两类:促铁死亡和抗铁死亡。我们发现,随着NRF2的改变,细胞内亚铁水平可能是决定细胞命运的关键因素。在TNBC细胞中亚铁浓度较高的情况下,PRMT5抑制NRF2/HMOX1通路并减缓亚铁的输入。此外,高PRMT5蛋白水平表明TNBC对免疫疗法具有较强的抵抗力,而PRMT5抑制剂可增强免疫疗法的治疗效果。结论我们的研究结果表明,PRMT5的激活可以调节铁代谢并驱动对铁死亡诱导剂和免疫疗法的抵抗。因此,PRMT5可以作为改变TNBC免疫抗性的靶标。

使用 CRISPR/Cas9 进行 T 细胞 Nrf2/Keap1 基因编辑和实验性肾脏缺血-再灌注损伤
目的:T 细胞在肾脏缺血再灌注损伤 (IRI) 中发挥病理生理作用,核因子红细胞 2 相关因子 2/kelch 样 ECH 相关蛋白 1 (Nrf2/Keap1) 通路调节 T 细胞反应。我们假设成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 9 (Cas9) 介导的 Keap1 敲除 (KO) 增强了 CD4+ T 细胞的 Nrf2 抗氧化潜力,而 Keap1 -KO CD4+ T 细胞免疫疗法可预防肾脏 IRI。结果:CD4+ T 细胞 Keap1-KO 导致 Nrf2 靶基因 NAD(P)H 醌脱氢酶 1、血红素加氧酶 1、谷氨酸-半胱氨酸连接酶催化亚基和谷氨酸-半胱氨酸连接酶修饰亚基显著增加。体外,Keap1-KO 细胞没有显示出衰竭迹象,在常氧条件下白细胞介素 2 (IL2) 和 IL6 水平显著降低,但在缺氧条件下干扰素 γ 水平升高。体内实验中,与接受未编辑对照 CD4+ T 细胞的小鼠相比,IRI 前过继转移 Keap1-KO CD4+ T 细胞可改善 T 细胞缺陷 nu/nu 小鼠的肾功能。与从对照肾脏中分离的未编辑 CD4+ T 细胞相比,IRI 后 24 小时从受体肾脏中分离的 Keap1-KO CD4+ T 细胞活性较低。创新:使用 CRISPR/Cas9 编辑小鼠 T 细胞中的 Nrf2/Keap1 通路是一种创新且有前景的免疫治疗方法,可用于治疗肾脏 IRI 以及其他实体器官 IRI。结论:CRISPR/Cas9 介导的 Keap1 -KO 增加了小鼠 CD4+ T 细胞中 Nrf2 调节的抗氧化基因表达,改变了对体外缺氧和体内肾脏 IRI 的反应。针对 T 细胞中 Nrf2/Keap1 通路的基因编辑是治疗免疫介导肾脏疾病的一种有前景的方法。抗氧化剂。氧化还原信号。38,959–973。

基于硅胶的发现,使用药效团筛查,分子对接和MD模拟研究的策略 神经元 - 胶体传播中的外泌体 一种基于模糊的逻辑计算方法,用于重新利用Covid-19 靶向缺氧诱导因子1α通过调节乳房can中的肿瘤微环境 制造介孔二氧化硅纳米颗粒,以靶向糖替尼向卵巢癌细胞靶向递送
摘要简介:这项研究的主要目的是确定开发有效KEAP1抑制剂的潜在潜在客户。方法:在当前的研究文章中,已采用了硅内方法来发现潜在的KEAP1抑制剂。3D-QSAR是使用具有IC 50的KEAP1抑制剂的Chembl数据库生成的。选择了最好的药理,以筛选三个不同的文库,即Asinex,Minimaybridge和锌。从数据库中筛选的分子通过可药物性规则和分子对接研究过滤。对接研究后获得的最佳结合分子通过二利方法对测定物理化学性质进行毒性进行。通过分子动态模拟,研究了最佳的命中以在KEAP1腔中进行稳定。结果:分别对不同数据库进行虚拟筛选,并获得了三个导线。这些铅分子asinex 508,minimaybridgehts_01719和锌0000952883在KEAP1腔中显示出最好的结合。铅的结合复合物的分子动态模拟支持对接分析。铅(Asinex 508,minimaybridgehts_01719和锌0000952883)在100 NS模拟的KEAP1结合腔中稳定,平均RMSD值分别为0.100、0.114和0.106 Nm。结论:这项研究提出了三个铅分子作为基于高吞吐量筛查,对接和MD模拟研究的潜在KEAP1抑制剂。这些HIT分子可用于进一步设计和开发KEAP1抑制剂。这项研究提供了用于发现新型KEAP1抑制剂的初步数据。它为药物化学家打开了新的途径,以探索针对KEAP1-NRF2途径的抗氧化剂刺激分子。

基于 PET/CT 的放射基因组学支持 KEAP1/NFE2L2 通路靶向治疗接受根治性放射治疗的非小细胞肺癌
对于肺癌患者,与手术相比,放射治疗会增加局部复发 (LR) 的风险,但毒性特征更佳。KEAP1/NFE2L2 突变状态 (Mut KEAP1/NFE2L2) 与接受放射治疗的患者的 LR 显着相关,但很少可用。使用非侵入性方式预测 Mut KEAP1/NFE2L2 有助于进一步个性化每种治疗策略。方法:基于 770 名患者的公共队列,首先使用连续基因表达水平开发模型 RNA (M-RNA) 来预测 Mut KEAP1/NFE2L2 ,从而得到二元输出。然后构建模型 PET/CT (M-PET/CT) 以使用 PET/CT 提取的放射组学特征来预测 M-RNA 二元输出。 M-PET/CT 在接受根治性容积调强弧形放射治疗的 151 名外部队列中得到了验证。每个模型都是使用多层感知器网络方法在单独的队列上构建、内部验证和评估的。结果:M-RNA 在测试队列中的 C 统计量为 0.82。在 101 名患者的训练队列中,保留的 M-PET/CT 的曲线下面积为 0.90(P,0.001)。将 20% 的概率阈值应用于测试队列后,M-PET/CT 的 C 统计量达到 0.7。由于患者根据 LR 风险显著分层,风险比为 2.61(P = 0.02),因此在容积调强弧形放射治疗队列中验证了相同的放射组学模型。结论:我们的方法可以使用 PET/CT 提取的放射组学特征预测 Mut KEAP1/NFE2L2,并有效地对接受放射治疗的外部队列中有 LR 风险的患者进行分类。

和 keap1b 基因敲除斑马鱼
Keap1 – Nrf2 通路是一种进化保守的机制,可保护细胞免受氧化应激和亲电试剂的侵害。在稳态条件下,Keap1 与 Nrf2 相互作用并导致其快速蛋白酶体降解,但当细胞暴露于氧化应激/亲电试剂时,Keap1 会感知它们,导致 Keap1 – Nrf2 相互作用不当和 Nrf2 稳定。因此,Keap1 被认为是 Nrf2 激活的“抑制剂”和“应激传感器”。有趣的是,鱼类和两栖动物有两种 Keap1(Keap1a 和 Keap1b),而哺乳动物、鸟类和爬行动物只有一种。系统发育分析表明,哺乳动物 Keap1 是鱼类 Keap1b 的直系同源物,而不是 Keap1a。在本研究中,我们使用斑马鱼遗传学研究了 Keap1a 和 Keap1b 之间的差异和相似之处。我们构建了 keap1a 和 keap1b 的斑马鱼基因敲除系。两种基因敲除系的纯合突变体均可存活且可育。在两种突变幼虫中,Nrf2 靶基因的基础表达和抗氧化活性均以 Nrf2 依赖的方式上调,表明 Keap1a 和 Keap1b 均可作为 Nrf2 抑制剂发挥作用。我们还分析了 Nrf2 激活剂萝卜硫素对这些突变体的影响,发现 keap1a- ,而非 keap1b- ,基因敲除幼虫对萝卜硫素有反应,表明两种 Keap1 的压力/化学感应能力不同。
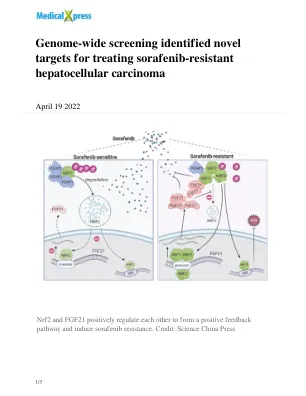
全基因组筛选确定了治疗索拉非尼耐药性肝细胞癌的新靶点
浙江大学医学院附属邵逸夫医院普通外科蔡秀军课题组领导的研究通过CRISPR/cas9系统对索拉非尼治疗下的肝癌细胞(HepG2)全基因组进行筛选,筛选出了在索拉非尼耐药中占主导地位的基因:KEAP1。KEAP1调控的下游分子Nrf2是细胞抵抗活性氧(ROS)的重要分子。本研究首先通过KEAP1/Nrf2基因编辑检测索拉非尼在肝癌细胞中的IC 50 等大量功能性实验,验证了KEAP1-Nrf2轴在索拉非尼耐药中的作用。本研究发现一种名为ML385的特异性Nrf2小分子抑制剂在体内和体外均能增强索拉非尼的杀伤作用。

将基因组生物标志物结合起来指导非小细胞肺癌Joris van de haar 1,2,3,Joanne M. Mankor 4,Karlijn Hummelink 5
摘要◥目的:在非小细胞肺癌(NSCLC)中指导免疫检查点阻断(ICB)治疗的STK11,KEAP1和EGFR改变的临床值仍然有争议,因为一些拟议的抗药性生物标志物显示出耐用的ICB反应。这种疾病迫切需要更多特定的组合生物标志物方法。ExperimentalDesign: Todevelopacombinatorialbiomarkerstra- tegy with increased speci fi city for ICB unresponsiveness in NSCLC, we performed a comprehensive analysis of 254 patients with NSCLC treated with ligand programmed death-ligand 1 (PD-L1) blockade monotherapy, including a discovery cohort of 75 patients subjected to whole-genome sequencing (WGS), and独立的验证队列,由169名患者进行肿瘤非正式面板序列。在低(<10 muts / mb)或高(≥10cuts / mb)肿瘤突变负担(TMB)的背景下,评估了STK11 / KEAP1 / EGFR改变的特定级别的ICB反应性。