XiaoMi-AI文件搜索系统
World File Search SystemLOXP

解锁 loxP 来追踪体内基因组编辑
摘要:CRISPR 相关蛋白(如 Cas9)的开发提高了基因组编辑的可及性和易用性。然而,需要额外的工具来量化和识别活体动物中成功的基因组编辑事件。我们开发了一种快速量化和监测活体动物中基因编辑活动的方法,该方法还有助于共聚焦显微镜和核苷酸水平分析。在这里,我们报告了一种新的 CRISPR“指纹识别”方法,用于激活小鼠中的荧光素酶和荧光蛋白作为基因编辑的功能。该系统基于我们之前的 cre 重组酶 (cre) 检测系统的经验,专为能够靶向 lox P 的 Cas 编辑器而设计,包括 SaCas9 和 ErCas12a 的 gRNA。这些 CRISPR 专门在 lox P 内切割,这种方法不同于以前靶向相邻终止序列的体内基因编辑活动检测技术。在这种传感器范例中,在肌肉或静脉内流体动力质粒注射后,在活体 cre 报告小鼠(FVB.129S6(B6)-Gt(ROSA)26Sortm1(Luc)Kael/J 和 Gt(ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo/J,本文中将称为 LSL-luciferase 和 mT/mG)中非侵入性地监测 CRISPR 活性,证明了其在两种不同器官系统中的实用性。通过共聚焦显微镜在特定组织的细胞水平上检查了相同的基因组编辑事件,以确定成功基因组编辑细胞的身份和频率。此外,SaCas9 诱导的靶向编辑效率与 cre 相当,证明了在整个动物中具有高效的传递和活性。这项研究建立了基因组编辑工具和模型,以非侵入性方式追踪体内 CRISPR 编辑并识别目标细胞。这种方法还使之前生成的数千种 lox P 动物模型中的任何一种都具有类似的实用性。

使用CRE/LOXP重组系统靶向基因整合到微藻的核基因组
抽象的遗传修饰的微藻被认为是生物能源和重组蛋白质产生的有用工具。然而,微藻核基因组中转基因的随机整合易受异源基因表达的基因沉默。在这里,我们试图使用CRE/ LOXP重组系统进行稳定的转基因表达,将靶向基因整合到雷目层的预定的核基因组位点中。我们构建了一个表达载体质粒编码报告基因(Zeocin耐药基因和绿色荧光蛋白基因; ZEO-2A-GFP)和突变的LOXP来产生创建者细胞。构建了编码IFNα-4的供体载体和抗性霉素的抗性基因,构造了相应突变的LOXP S,并与CRE表达载体一起构建并引入创始人细胞。通过计算抗霉素抗性菌落的数量来确定供体载体与CRE表达载体的最佳比率。对于已建立的克隆,使用各种特定引物集通过基因组PCR确认了靶向积分。供体载体中的靶基因可以使用CRE/ LOXP系统整合到Reinhardtii的预期基因组位点中。rt-PCR表明,IFNα-4在测试的五个独立的转基因细胞系中表达。该结果表明,基于CRE的细胞工程是一种产生表达外源基因的智能微藻的有前途的方法。


使用I-Gonad方法
CRE/ LOXP系统是体内基因功能研究的强大工具。CRE重组酶的调节表达以空间和时间控制的方式介导了遗传因素的精确缺失。尽管该系统具有鲁棒性,但仍需要大量精力为每个感兴趣的单个基因创建有条件的敲除模型,在这些基因中,必须同时将两个LOXP位点插入顺式。当前的工作涉及劳动密集型胚胎(ES)细胞 - 基于基因靶向小鼠胚胎的基因和乏味的微观畅通。该工作流的复杂性构成了技术挑战,因此限制了有条件遗传学的更广泛应用。在这里,我们通过将CRISPR供体的独特设计与新的Oviduct电穿孔技术I I -Gonad集成了一种替代方法来生成鼠标LOXP等位基因。显示了这种方法的潜力和简单性,我们在一次尝试中创建了五个基因的FloxErx等位基因,其成本相对较低,设备设置最少。除了条件等位基因外,还获得了本构敲除等位基因作为这些实验的副产品。因此,I -Gonad的更广泛应用可能会使用新型鼠模型促进基因功能研究。

利用 i-GONAD 进行多次连续基因组编辑和育种富集有助于生产转基因小鼠
摘要:转基因 (GM) 小鼠是生物医学研究中必不可少的工具。传统的转基因小鼠生成方法成本高昂,需要专门的人员和设备。使用成簇的规律间隔短回文重复序列 (CRISPR) 结合改进的输卵管核酸递送基因组编辑 (i-GONAD) 大大提高了在研究实验室中生产转基因小鼠的可行性。然而,由于 C57BL/6 (B6) 等近交系小鼠的生育能力低且胚胎脆弱,对其进行基因改造仍然具有挑战性。我们在尝试优化 i-GONAD 的同时,已在 B6 背景下成功生成了多种新型转基因小鼠品系。我们发现 i-GONAD 减少了超排卵怀孕雌性的产仔数,但不会影响怀孕率。自然交配或低激素剂量不会增加超排卵 B6 雌性中观察到的低生育率。然而,饮食丰富对怀孕成功有积极影响。我们还通过将接受 i-GONAD 治疗的怀孕 B6 雌性与同步怀孕的 FVB/NJ 伴母共同饲养来优化繁殖条件,以提高幼崽的存活率。因此,通过丰富的饮食和与生育能力强的雌性(如 FVB/NJ)共同抚养幼崽,增加了转基因小鼠的产生。在本研究中,我们使用 CRISPR/Cas 系统同时或连续靶向单个和多个基因座,产生了 16 只转基因小鼠。我们还比较了使用不同方法插入 LoxP 以产生条件性敲除小鼠的同源定向修复效率。我们发现,两步连续 LoxP 插入(其中每个 LoxP 序列在不同的 i-GONAD 程序中单独插入)是一种低风险、高效的产生 floxed 小鼠的方法。

基因组编辑项目设计的一般准则和实用技巧.docx
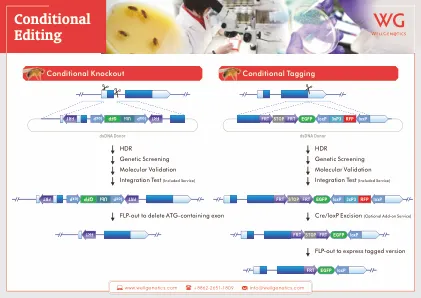
方法。第一种方法是将含有 loxP 位点的 ssODN 引入目标外显子两侧的 5' 和 3' 位点。这是通过使用 2 个 sgRNA 完成的。第二种方法使用含有 2 个 loxP 位点的 lssDNA 模板,这 2 个 loxP 位点位于目标 DNA 序列两侧。这种方法使用了 2 个 sgRNA。B. UTSW 转基因核心提供给您的试剂:您向核心支付的 CRISPR 服务费用包括我们用于完成您的项目的 IDT Sp. Cas9 蛋白的费用。如果您的项目涉及使用不同的编辑酶(如 Cas12 或 Cpf),请联系核心工作人员更详细地讨论该项目。C. 了解您的基因的重要细节:使用基因组浏览器(如 NCBI、UCSC 或 ENSEMBl)收集有关您的目标基因的相关信息。这包括任何替代转录本、外显子的数量和重要性、位于特定内含子中的调控基序以及编码序列等细节,以便成功设计 sgRNA 和供体 DNA 模板。使用 MGI- 小鼠基因组信息学来确定是否存在与靶基因敲除相关的已知表型非常重要。了解基因的 KO 是否可能导致致命表型(无论是胚胎还是出生后早期)尤为重要。了解并将此信息传达给核心人员将使我们能够修改用于生成小鼠突变株的条件,以便我们主要创建 KO 等位基因杂合的小鼠。D. sgRNA 的设计:注射受精卵中发生的基因组编辑的效率在很大程度上取决于针对靶标的 sgRNA 的正确设计。此设计的关键组成部分包括:

技术和策略测序
Pierce等,Proc。 natl。 学院。 ski.u.s.a. (1992)89,2056-2060•2 LOXP重组站点由Ricombinase CRE识别•PAC网站•PAC网站(162 bp),用于混合病毒颗粒中的重组分子•“填充”的“填充器”片段的“填充”(填充11 kb的adenovirus dna plapies plapies plase prastic•gene prast)复制品岩性的大量副本(在LAC启动子下)•SACB编码为Levan Sucrasi的SACB,催化蔗糖水解的酶。 在大肠杆菌中表达时,神圣的酶会产生levano,该酶积聚在周质空间中,对细胞的致命作用Pierce等,Proc。natl。学院。ski.u.s.a.(1992)89,2056-2060•2 LOXP重组站点由Ricombinase CRE识别•PAC网站•PAC网站(162 bp),用于混合病毒颗粒中的重组分子•“填充”的“填充器”片段的“填充”(填充11 kb的adenovirus dna plapies plapies plase prastic•gene prast)复制品岩性的大量副本(在LAC启动子下)•SACB编码为Levan Sucrasi的SACB,催化蔗糖水解的酶。在大肠杆菌中表达时,神圣的酶会产生levano,该酶积聚在周质空间中,对细胞的致命作用
梅兰妮·古利
威康基金会桑格研究所 – 硕士论文 使用 prime editing 随机化基因调控区域 通过分析全基因组染色质数据集并将研究结果与文献检索相结合,确定了有趣的增强子区域。 使用 CRISPR prime editing 将多个重组酶识别序列插入这些基因的增强子簇中。 创建了具有稳定 prime editor 表达的细胞系,可实现 loxP 位点 80% 以上的插入效率(这些细胞系现在被实验室中的其他人广泛使用)。 在实验室中建立了具有 Cas9 富集的靶向牛津纳米孔测序。 与帝国理工学院的一个团队合作学习该方法。 在 2022 年国际哺乳动物合成生物学会议上以海报形式展示了我的工作。 2021 年 11 月 12 日,法国斯特拉斯堡

IHC-2023-ABSTRACTs.pdfIHC-2023-ABSTRACTs.pdf
蛋白质,i)CLR,II)ramp1和iii)CGRP受体成分蛋白(RCP),这是通过G A S途径发出信号所需的细胞内周围膜蛋白。虽然当前的疗法集中于拮抗CGRP或CLR/RAMP1复合物,但我们投资了CGRP-RCP的损失会导致运动引起的恶心疼痛。方法:我们已经生成了CRCP基因第二外显子的LOXP条件的小鼠。然后,我们用Nestin-creer小鼠将这些CRCP-LoxP小鼠越过,导致他莫昔芬诱导后的神经元RCP表达障碍的小鼠。他莫昔芬诱导的CRCP-loxp小鼠未表达CRE作为对照。在他莫昔芬诱导后,两组小鼠在运动引起的恶心的测定中进行了测试。Mice were assessed after intraperitoneal (IP) injections of: i) vehicle-PBS, ii) CGRP (0.1 mg/kg), or iii) CGRP (0.1 mg/kg) co-delivered with either olcegepant (1.0 mg/kg-CGRP-receptor antago- nist) or rizatriptan (1.0 mg/kg-selective serotonin receptor激动剂)。结果:我们观察到CGRP增加了运动诱发的恶心,而Olcegepant(但不是Rizatriptan)能够减轻CGRP在他莫昔芬治疗的对照小鼠(N¼11F/11M)中的影响。有趣的是,floxed cgrp-rcp null( - / - )小鼠的反应类似于对照对照,但olcegepant和rizatriptan均未减轻CGRP的效果(n¼11f/11m);表明不存在CGRP-RCP会使受体对拮抗剂Olcepant没有反应。致谢:这项研究得到了NIH R01 DC017261(AEL)和罗切斯特大学旋转格兰特(IMD)的支持。披露利息:无声明结论:总而言之,我们的发现表明:i)系统CGRP增加了缺乏神经元CGRP-RCP的对照和小鼠运动引起的运动引起的恶心; ii)细胞内CGRP-RCP的丧失会使CLR/ RAMP1复合物对OlcePant不敏感,从而显示出CGRP受体处的反抗偏见。