XiaoMi-AI文件搜索系统
World File Search SystemMSTN
利用 CRISPR/Cas9 破坏大黄鱼 (Larimichthys crocea) 中的 mstn 基因
mstnab +/− 鲤鱼的体长显著增加(Shahi et al.,2022),为经济鱼类的养殖产量提供了有希望的方向。因此,我们选择大黄鱼的mstnb作为靶基因。经检测,我们设计的针对外显子1中编码序列的8个靶标中的两个sgRNA是有效的(图1b)。与野生型鱼中的序列相比,检测到了5个缺失突变,包括同时发生的12 bp、28 bp、36 bp、83 bp和97 bp缺失(图2c)。与单个gRNA微注射(Shahi et al.,2022,Tao et al.,2021,Zhang et al.,2020b)不同,同时注射多个gRNA可能诱导两个靶位点之间更大片段的缺失(图2c)。该方法也被考虑并应用于斑马鱼视网膜疾病模型研究中,采用基于CRISPR/Cas9系统的更快速有效的策略

由 CRISPR-Cas9 系统指导的突变绵羊双肌表型
肌生长抑制素 (MSTN) 是一种众所周知的肌肉生长负调节剂。由 MSTN 自然功能丧失突变引起的双肌羊具有非常强的骨骼肌。在这项研究中,我们的结果表明,通过使用 Cas9 技术特异性靶向外显子 1 位点,成功生成了 MSTN 突变羊。我们研究中的 MSTN 敲除羊的肌肉显著增加,就像双肌表型一样。我们的研究表明,将 Cas9:sgRNA 直接注射到受精卵中可广泛用于在大型家畜中产生基因敲除。值得注意的是,根据我们的研究结果,绵羊可以加入到现在越来越实用的基因组编辑物种名单中。MSTN 突变羊的生成对于当地绵羊品种的遗传改良以及将绵羊用作大型动物医学研究的模型具有重要意义。
Joonbum Lee1,2、Dong-Hwan Kim1 和 Kichoon ...
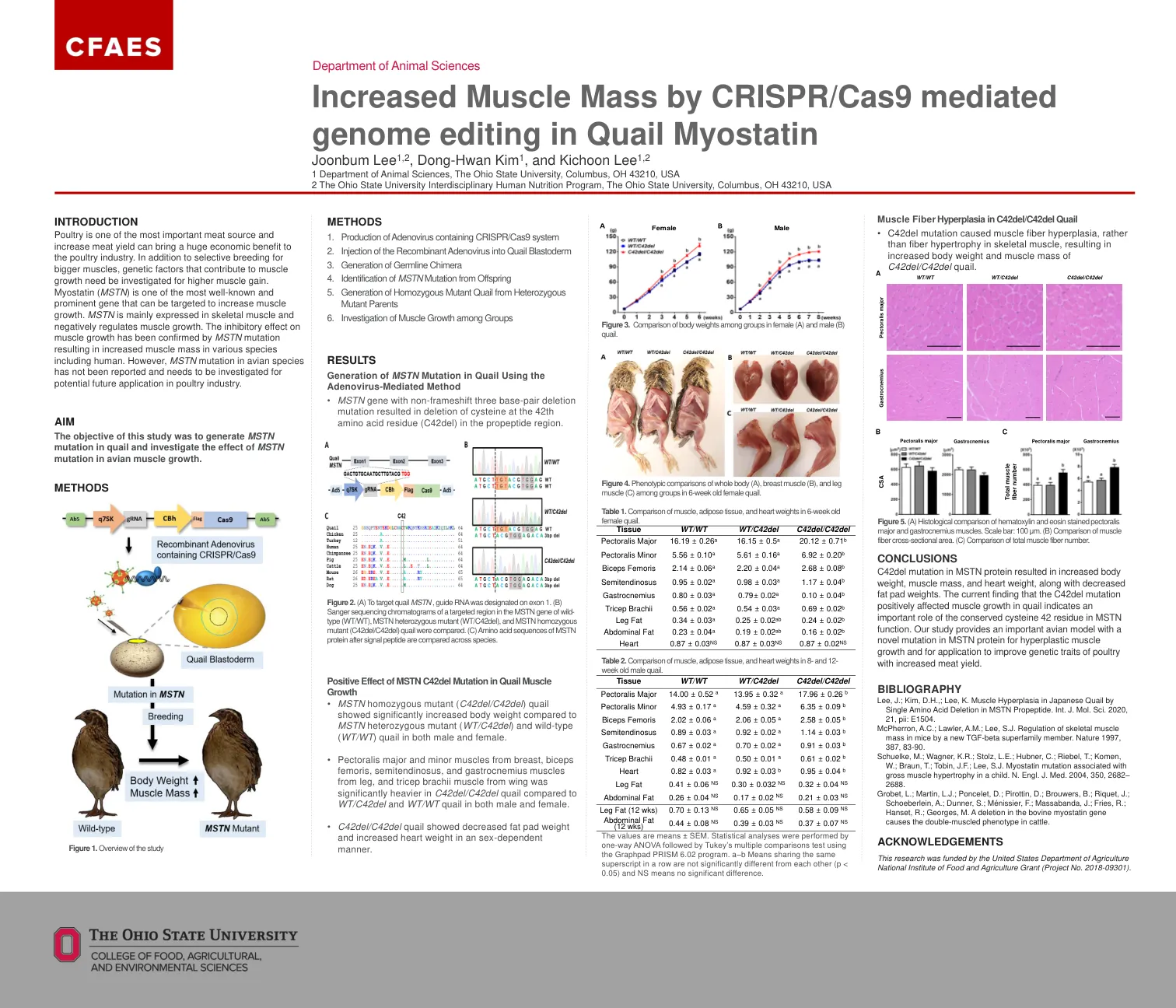
引言家禽是最重要的肉类来源之一,提高肉类产量可为家禽业带来巨大的经济效益。除了选择性育种以获得更大的肌肉外,还需要研究有助于肌肉生长的遗传因素以获得更高的肌肉增益。肌生长抑制素 (MSTN) 是最著名和最突出的基因之一,可以作为增加肌肉生长的靶点。MSTN 主要在骨骼肌中表达,并对肌肉生长有负向调节作用。MSTN 突变已证实对肌肉生长有抑制作用,导致包括人类在内的各种物种的肌肉质量增加。然而,尚未见鸟类 MSTN 突变的报道,需要进行研究以期未来将其应用于家禽业。

GDF11 和 GDF8 之间不同的氨基酸功能替代会影响骨骼发育和骨骼肌
生长分化因子 11 (GDF11) 和 GDF8 (MSTN) 是密切相关的 TGF- β 家族蛋白,它们与几乎相同的信号受体和拮抗剂相互作用。然而,GDF11 在体外和体内似乎比 GDF8 更有效地激活 SMAD2/3。配体具有不同的结构特性,将独特的 GDF11 氨基酸替换到 GDF8 中可增强所得嵌合 GDF8 的活性。我们通过基因改造 GDF11 和 GDF8 的成熟信号结构域,研究了它们在体内可能不同的内源性活性。将 GDF8 完全重新编码为 GDF11 会产生缺乏 GDF8 的小鼠,其 GDF11 水平比正常水平高出约 50 倍,肌肉质量略有下降,但对健康或生存没有明显的负面影响。将 GDF11 指尖区域的两个特定氨基酸替换为相应的 GDF8 残基,可导致产前轴向骨骼转变,与 Gdf11 缺陷小鼠一致,且骨骼或心肌发育或体内平衡没有明显紊乱。这些实验揭示了体内 GDF11 和 GDF8 成熟结构域之间的独特特征,并确定了早期骨骼发育对 GDF11 的特定要求。

牛和猪基因组中可能致病变异的证据和定位
摘要背景:本文回顾了当代猪和牛参考基因组中已发表的潜在致病变异的定位及其因果关系的证据。尽管从基因图谱和全基因组关联研究中鉴定致病变异本身就很困难,但动物遗传学研究人员已经针对几种与牲畜育种相关的性状提出了推定的致病变异。结果:为了进行这篇综述,我们阅读了支持牛和猪的 13 个基因(ABCG2、DGAT1、GHR、IGF2、MC4R、MSTN、NR6A1、PHGK1、PRKAG3、PLRL、RYR1、SYNGR2 和 VRTN)存在潜在致病变异的文献,并将它们定位在当代参考基因组中。我们审查了它们之间的因果关系的证据,旨在将基因座、拟议的致病基因和拟议的致病变异的证据区分开来,并报告在牛或猪基因组中定位序列变异所需的生物信息学搜索和策略。结论:总而言之,通常有很好的证据表明基因座水平存在关联,八个基因座存在特定致病基因的证据,六个基因座存在特定致病变异的一些实验证据。我们建议报告新的潜在致病变异的研究人员使用参考坐标系统,显示本地序列上下文,并将变异提交到存储库。

CRISPR/Cas9 介导的猪受精卵基因组编辑前的透明带处理
Abkowitz, JL、Persik, MT、Shelton, GH、Ott, RL、Kiklevich, JV、Catlin, SN 和 Guttorp, P. (1995)。大型动物造血干细胞的行为。美国国家科学院院刊,92 (6),2031–2035。https://doi.org/10.1073/pnas.92.6.2031 Brinkman, EK、Kousholt, AN、Harmsen, T.、Leemans, C.、Chen, T.、Jonkers, J. 和 Van Steensel, B. (2018)。模板引导的 CRISPR/Cas9 编辑的简易量化。核酸研究,46 (10),e58。 https://doi.org/10.1093/nar/gky164 Le, QA, Hirata, M., Nguyen, NT, Takebayashi, K., Wittayarat, M., Sato, Y., Namula, Z., Nii, M., Tanihara, F., & Otoi, T. (2020)。使用不同浓度的 Cas9 蛋白和靶向肌肉生长抑制素 (MSTN) 基因的 gRNA 进行电穿孔处理对猪受精卵发育和基因编辑的影响。动物科学杂志,91 (1),e13386。 https://doi.org/10.1111/asj.13386 Li, R.、Liu, Y.、Pedersen, HS、Kragh, PM 和 Callesen, H. (2013)。猪单性生殖胚胎去除透明带后的发育和质量。Theriogenology,80 (1),58–64。https://doi.org/10.1016/j.theriogenology.2013.03.009 Meurens, F., Summerfield, A., Nauwynck, H., Saif, L., & Gerdts, V. (2012)。猪:人类传染病的模型。微生物学趋势,20 (1),50–57。Nishio, K., Tanihara, F., Nguyen, T.-V., Kunihara, T., Nii, M., Hirata, M., Takemoto, T., & Otoi, T. (2018)。电穿孔过程中电压强度对体外生产的猪胚胎发育和质量的影响。家畜繁殖,53 (2),313–318。https://doi. org/10.1111/rda.13106 Peng, H., Wu, Y., & Zhang, Y. (2012)。通过电穿孔将 DNA 和吗啉代诺西酮有效递送到小鼠植入前胚胎中。PLoS One,7 (8),e43748。https://doi.org/10.1371/journal.pone.0043748 Peura, TT, & Vajta, G. (2003)。绵羊和牛核移植中现有方法与新方法的比较。克隆干细胞,5 (4),257–277。 https://doi.org/10.1089/153623003772032772 Qin, W., Dion, SL, Kutny, PM, Zhang, Y., Cheng, AW, Jillette, NL, Malhotra, A., Geurts, AM, Chen, Y.-G., & Wang, H. (2015). 通过合子电穿孔核酸酶在小鼠中实现高效的 CRISPR/Cas9 介导基因组编辑。遗传学,200 (2), 423–430。 https://doi.org/10.1534/ Genetics.115.176594 Remy, S., Chenouard, V., Tesson, L., Usal, C., Ménoret, S., Brusselle, L., Hes- lan, J.-M., Nguyen, TH, Bellien, J., Merot, J., De Cian, A., Giovannangeli, C., Concordet, J.-P., &Anegon, I. (2017). 通过使用电穿孔将 CRISPR/Cas9 蛋白和供体 DNA 递送到完整受精卵中来生成基因编辑大鼠。科学报告,7 (1),16554。https://doi.org/10。 1038/s41598-017-16328-y Tanihara, F.、Hirata, M.、Nguyen, NT、Sawamoto, O.、Kikuchi, T.、Doi, M. 和 Otoi, T. (2020)。通过将 CRISPR/Cas9 系统电穿孔到体外受精的受精卵中有效生成 GGTA1 缺陷猪。BMC Biotechnology,20 (1),40。https://doi.org/10.1186/s12896-020-00638-7