XiaoMi-AI文件搜索系统
World File Search SystemMTORC1

携带EGFR突变的小细胞肺癌患者
抽象背景二甲双胍(MET)是2型糖尿病的一线治疗方法,在治疗各种疾病(例如心血管疾病,神经退行性疾病,癌症和衰老)中起着有效的作用。然而,依赖MET的抗肿瘤免疫的潜在机制仍有待阐明。方法Mitotempo是线粒体超氧化物的清道夫,废除了MET的抗肿瘤作用,但没有消除抗编程细胞死亡(PD-1)抗体(AB)治疗。因此,我们研究了遇到的抗肿瘤效应的机制。葡萄糖转运蛋白(GLUT)-1,线粒体活性氧(MTROS),干扰素(IFN)-γ,Ki67,自噬标记,NF-E2相关因子2(NRF2)的激活标记和乳腺脂蛋白复合物1(Mammaycin Confictrc 1(Mimamalian Infilc)1(mmAMAMAYCINC)1(MIMMAYCIN COFFILC 1)通过流式细胞仪分析检查淋巴细胞(CD8TIT)。此外,还使用有条件的NRF2和p62小鼠来检测这些标记,并监测体内肿瘤生长。对CD8TIT和肿瘤细胞进行 RNA测序。 黑色素瘤细胞含有IFN-γ受体(IFNγR)细胞质域缺失突变体过表达,并用于使用Seahorse Flux分析仪来表征这些肿瘤细胞的代谢谱。 结果MET给药可提高MTROS和细胞表面Glut-1,从而导致CD8TIT中的IFN-γ产生。 mtros以糖酵解依赖性方式激活NRF2,从而诱导自噬,谷氨酰胺溶解,mTORC1和p62/sqSTM1的激活。RNA测序。黑色素瘤细胞含有IFN-γ受体(IFNγR)细胞质域缺失突变体过表达,并用于使用Seahorse Flux分析仪来表征这些肿瘤细胞的代谢谱。结果MET给药可提高MTROS和细胞表面Glut-1,从而导致CD8TIT中的IFN-γ产生。mtros以糖酵解依赖性方式激活NRF2,从而诱导自噬,谷氨酰胺溶解,mTORC1和p62/sqSTM1的激活。在丝氨酸351(p-p62(S351))上,p62的MTORC1依赖性磷酸化也参与了NRF2的激活。CD8TIT中NRF2的条件缺失消除了MTORC1激活和抗肿瘤免疫。 与抗PD-1 AB的作用协同作用,满足CD8TIT增殖和IFN-γ分泌,从而导致肿瘤细胞中糖酵解和氧化磷酸化的降低。 因此,在CD8TIT中,GLUT-1以及活化的树突状细胞的扩展升高。 此外,缺乏IFNγR信号的肿瘤细胞消除了CD8TIT的IFN-γ的产生和增殖。CD8TIT中NRF2的条件缺失消除了MTORC1激活和抗肿瘤免疫。与抗PD-1 AB的作用协同作用,满足CD8TIT增殖和IFN-γ分泌,从而导致肿瘤细胞中糖酵解和氧化磷酸化的降低。因此,在CD8TIT中,GLUT-1以及活化的树突状细胞的扩展升高。此外,缺乏IFNγR信号的肿瘤细胞消除了CD8TIT的IFN-γ的产生和增殖。

mTOR 通路在肝癌中的作用
原发性肝癌,包括肝细胞癌 (HCC) 和肝内胆管癌 (iCCA),是全球范围内致死率极高且常见的肿瘤,且几乎没有有效的治疗方法。哺乳动物雷帕霉素靶蛋白 (mTOR) 复合物是细胞生长和代谢的中心调节器,它能够整合来自氨基酸、营养物质和细胞外信号的输入。mTOR 蛋白被整合到两个不同的复合物中:哺乳动物雷帕霉素靶蛋白复合物 1 (mTORC1) 和哺乳动物雷帕霉素靶蛋白复合物 2 (mTORC2)。具体而言,mTORC1 调节蛋白质合成、葡萄糖和脂质代谢以及自噬,而 mTORC2 通过调节 AGC 丝氨酸/苏氨酸激酶家族,尤其是 AKT(蛋白激酶 B)蛋白来促进肝脏肿瘤发生。在人类 HCC 和 iCCA 样本中,基因组学分析揭示了 mTOR 复合物频繁失调的情况。体外和体内研究均已证明 mTORC1 和 mTORC2 在肝肿瘤发展和进展中起着关键作用。第一代 mTOR 抑制剂在肝肿瘤治疗中的有效性已得到评估,但结果并不令人满意。当前的研究工作致力于生成更有效的 mTOR 抑制剂并确定用于患者选择和新型联合疗法的生物标志物。在这里,我们全面回顾了导致肝癌中失调的 mTOR 信号级联的机制、mTOR 通路促进 HCC 和 iCCA 分子发病机制的机制、治疗策略以及有效抑制肝癌治疗中的 mTOR 所需克服的挑战。总之:失调的 mTOR 信号对 HCC 和 iCCA 分子发病机制有显著贡献。mTOR 抑制剂(可能与其他药物联合使用)可能对人类肝肿瘤的亚群有效。

缓释雷帕霉素可阻止亚临床猫肥厚型心肌病左心室肥厚的进展:RAPACAT 的结果
雷帕霉素,又称西罗莫司,是一种大环内酯类药物,是雷帕霉素机制靶点(以前称为哺乳动物靶点)的抑制剂,有望成为治疗猫 HCM 的新型疗法。5–10 mTOR 是一种非典型丝氨酸/苏氨酸蛋白激酶,可与多种蛋白质结合形成 2 种多蛋白复合物中的 1 种:mTOR 复合物 1 (mTORC1) 和 mTOR 复合物 2 (mTORC2)。5,11,12 这两种复合物在促进胚胎发生和出生后心血管健康方面发挥着不同的作用,具有不同的上游和下游靶点。mTORC1 的激活可促进合成代谢过程,从而增加蛋白质和脂质的合成,下调分解代谢过程以减少自噬(细胞清除受损蛋白质和细胞器的能力),并在响应机械超负荷发出适应性心脏重塑信号方面发挥重要作用。 5 尽管尚未完全了解,但 mTORC2 在调节葡萄糖和脂质代谢中发挥作用,并促进心肌细胞存活、细胞骨架组织和适当的细胞极性。

MG53与2型糖尿病,血糖控制和糖尿病并发症的存在的关联
结节性硬化症复合物(TSC)是一种令人衰弱的发育障碍,其特征是多种临床表现。TSC是由TSC1或TSC2基因中的突变引起的,该突变分别编码了Hamartin/tuberin蛋白。这些蛋白质充当异二聚体,对雷帕霉素复合物1(MTORC1)的机械靶标进行负调节。TSC研究集中在MTORC1(一个关键信号中心)对调节包括代谢,细胞生长,翻译和神经发生在内的各种细胞过程的影响。然而,TSC2的非典型功能尚未得到很好的研究,并且对影响这些功能的突变的潜在潜在的生物学机制尚不清楚。我们观察到TSC2突变体IPSC中的异常多极有丝分裂分裂。多极表型不受抑制剂雷帕霉素治疗的有意义的影响。我们进一步观察到TSC2突变形式的主要负活性在产生多极分裂表型中。这些数据扩大了TSC2功能和病理生理学的知识,这将与TSC患者的未来治疗高度相关。

针对 S6K1 的化合物可阻碍脂肪量扩张并减轻饮食引起的肝脂肪变性
简介 肝脏和脂肪组织控制着体内脂质稳态。长期食用含有大量脂肪的饮食时,这些器官的相互功能障碍可能会加剧与肥胖相关的代谢紊乱 (1)。其中,血脂异常(包括高甘油三酯血症和高胆固醇血症)是肥胖相关代谢失衡的共同特征,可能引发一系列并发症,即所谓的代谢综合征 (2)。此外,肝脏脂肪变性是脂质稳态紊乱的关键致病因素,可加速动脉粥样硬化,并使血脂异常处于肥胖与心血管和代谢疾病风险的交汇点 (3–5)。因此,一种能降低脂肪量膨胀并改善肝脏脂质处理、预防肝脂肪变性和血脂异常的药理学化合物将为治疗与肥胖表型相关的代谢综合征带来重大进展。核糖体蛋白 S6 激酶 1 (S6K1) 在哺乳动物雷帕霉素靶蛋白复合物 1 (mTORC1) 下游起作用,后者控制对激素和有丝分裂原的反应,还协调细胞对营养物质和能量输入的反应 (6)。S6K1 的激活由一系列有序的构象变化和磷酸化步骤介导,其中 mTORC1 对 T389 的磷酸化为磷酸肌醇依赖性激酶 1 (PDK1) 创造了一个对接位点,从而允许 T229 磷酸化 (7)。

mTOR 信号传导:癌症、心血管疾病、糖尿病和衰老的新见解
雷帕霉素的机制/哺乳动物靶点 (mTOR) 是磷酸肌醇 3-激酶 (PI3K) 相关激酶家族的成员,它整合细胞内和环境因素,协调多种细胞/组织功能,如细胞生长、增殖、代谢、自噬、细胞凋亡、寿命、蛋白质/脂质/核苷酸合成以及组织再生和修复 [1]。尽管 mTOR 信号对于正常的细胞稳态至关重要,但 mTOR 的异常激活可能与多种病理结果相关,包括不同类型的癌症、代谢/心血管/肺部疾病和神经退行性疾病 [2]。考虑到 mTOR 信号转导的病理生理重要性,我们在本期特刊中收集了评论文章、原创研究文章和简短通讯,以深入了解不同疾病中的 mTOR 信号网络,从而开发新的 mTOR 靶向治疗方法。 mTOR 是两个结构和功能不同的蛋白质复合物的核心成分,即 mTOR 复合物 1(mTORC1)和 mTOR 复合物 2(mTORC2)。mTORC1 整合各种刺激和信号网络来促进细胞合成代谢,但它通过调节自噬过程的每个步骤(包括诱导、成核、延长和双膜自噬体的形成,自噬溶酶体的形成以及自噬体隔离底物的回收)来抑制自噬等分解代谢过程[3]。肌肉中 mTORC1-自噬轴的失调可导致多种肌肉疾病的发展。Han 等人强调了平衡 mTORC1 和自噬在能量产生/消耗和大分子周转过程中的重要作用,对于维持骨骼肌的生理状态[4]。他们还讨论了通过恢复 mTORC1 和自噬之间的平衡来减轻两种肌肉疾病(癌症恶病质和肌肉减少症)进展的潜在治疗选择。PI3K/mTOR 信号通路在调节细胞稳态中起着关键作用;因此,信号失调通常与衰老和与年龄相关的病理有关,包括癌症、心血管疾病和糖尿病等。因此,了解这种高度非线性的系统通路(涉及复杂的调节机制和与邻近通路的串扰)对于推进生物学和开发新的治疗方法是必不可少的。Ghomlaghi 等人利用文献中可用的计算和实验研究,对 PI3K/mTOR 信号的复杂动态机制网络进行了很好的概述,强调了它与其他主要信号通路的相互作用/相互依赖性、竞争性抑制和表观遗传改变特性,以及相关的相互关联的正/负调节回路(反馈/前馈机制)[ 5 ]。在本文中,作者强调了使用计算模型来研究这种复杂的网络
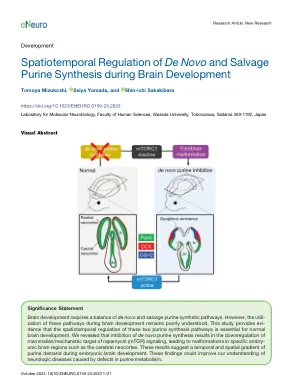
大脑发育过程中从头和挽救嘌呤合成的时空调节
嘌呤的水平,维持真核细胞体内平衡的必需分子,受到从头和救助合成途径的坐标的调节。在胚胎中枢神经系统(CNS)中,从头途径对于满足神经茎/促生细胞(NSPC)主动扩散的要求被认为至关重要。但是,在中枢神经系统开发期间,这两种途径如何平衡或分别使用。在这项研究中,我们显示了途径利用率的动态变化,并且在胚胎阶段和产后 - 成年小鼠脑的拯救途径上更依赖于从头途径。各种嘌呤合成抑制剂在体外的药理作用以及嘌呤合成酶的表达概况表明,胚胎大脑中的NSPC主要使用从头途径。同时,小脑中的NSPC同时需要从头和打捞途径。在从头抑制剂的体内给药导致前脑皮质区域严重下降症,表明沿胚胎大脑的前后轴沿着嘌呤的嘌呤需求梯度,而背侧前脑的皮质区域比腹膜或腹膜较高的嘌呤需求更高。 这种新皮层的组织学缺陷伴随着雷帕霉素复合物1(MTORC1)/核糖体蛋白S6激酶(S6K)/S6信号传导壳的强烈下调,这是一种至关重要的途径,用于细胞代谢,生长和生存。在从头抑制剂的体内给药导致前脑皮质区域严重下降症,表明沿胚胎大脑的前后轴沿着嘌呤的嘌呤需求梯度,而背侧前脑的皮质区域比腹膜或腹膜较高的嘌呤需求更高。这种新皮层的组织学缺陷伴随着雷帕霉素复合物1(MTORC1)/核糖体蛋白S6激酶(S6K)/S6信号传导壳的强烈下调,这是一种至关重要的途径,用于细胞代谢,生长和生存。这些发现表明,嘌呤途径对MTORC1信号传导和适当脑发育的时空调节的重要性。

高脂饮食通过诱导肠道菌群介导的亮氨酸产生和PMN-MDSC分化来促进癌症的进展
这项研究表明,高脂肪饮食(HFD) - 相关的肠道菌群通过激活乳腺癌和黑色素瘤模型中的多形核骨髓细胞(PMN -MDSC)产生来促进叶酸以促进癌症的进展。HFD微生物群通过触发髓样祖细胞中的MTORC1信号通路来释放丰富的亮氨酸,从而诱导PMN -MDSC产生。因此,在HFD驱动的肿瘤发生中建立了“肠道骨髓 - 肿瘤”轴。我们的观察结果还表明,DeLfovibrio属富含超重/肥胖患者的粪便,并与肿瘤生长,粪便和PMN -MDSC水平呈正相关,这表明desulfofibibio属是DeSulfibibibio属的肠菌菌群的关键成分,导致了癌症的癌症进展。

针对卵巢癌中的 PI3K/AKT/mTOR/NFκB 轴
大多数卵巢癌病例,无论亚型如何 [8]。PIK3CA 突变被认为是驱动突变,为高级别浆液性癌 (HGSC) 提供转化优势 [9]。多变量生存分析显示,PI3K 蛋白表达与晚期 HGSC 的较差生存率相关 [10]。此外,一些研究表明,PI3K 通路中的突变率,尤其是 AKT 和 p70S6K 中的突变率,包括错义突变和扩增,与较高的化学耐药率相关 [11,12]。化学增敏可以通过下调 PI3K 和/或其下游效应物 AKT 和 mTORC1 来实现 [13-15]。PI3K 在 OvCa 中的活性增加及其作为几种促癌通路的枢纽的作用,解释了其在癌症进展中的许多影响,包括致癌转化、

调节肠道干细胞体内平衡的营养代谢
图2饮食模式调节肠道干细胞(ISC)功能。(a)禁食,快速恢复和卡路里限制。禁食通过启动脂肪酸氧化(FAO)程序来增强ISC功能,并取决于肉碱棕榈酰转移酶1A(CPT1A)。其他调节器(例如PRDM16和HNF4A/G)也通过调节粮农组织来调节ISC。快速恢复后刺激MTORC1并通过多胺代谢程序激活蛋白质合成。结果,ISC增殖和肿瘤发生都升高。在卡路里限制期(CR)期间,由于雷帕霉素复合物1(MTORC1)的机理靶标降低,paneth细胞旁分泌因子循环ADP核糖(CADPR)。CADPR进入ISC,并通过SIRT/MTORC1-S6K1信号传导促进ISC和Paneth细胞的增殖。CR还增强了储备ISC中的DNA损伤性,从而保留了再生能力。(b)高脂,高脂/高糖和生酮饮食。高脂饮食(HFD)通过过氧化物酶体增殖物 - 活化受体δ(PPARδ)和LXR/FXR信号传导激活β-蛋白酶靶基因,从而促进ISC增殖。此外,PPARδ使祖细胞能够恢复干细胞特征,从而促进肿瘤发生。高脂/高糖饮食(HFHSD)通过激活固醇调节性元件结合蛋白1(SREBP1,用于脂肪酸合成),PPARγ信号传导和胰岛素受体-FR-FR-FR-AKT途径来诱导粘膜变化和肠道疾病。酮体衍生自生酮饮食(KTD)或禁食会影响ISC茎和通过3-羟基-3-甲基戊二核酸COA合成酶2(HMGCS2)-Class-Class-Class 1组蛋白脱乙酰基酶(HDAC) - NOTCH信号的分化。