XiaoMi-AI文件搜索系统
World File Search SystemMeis1
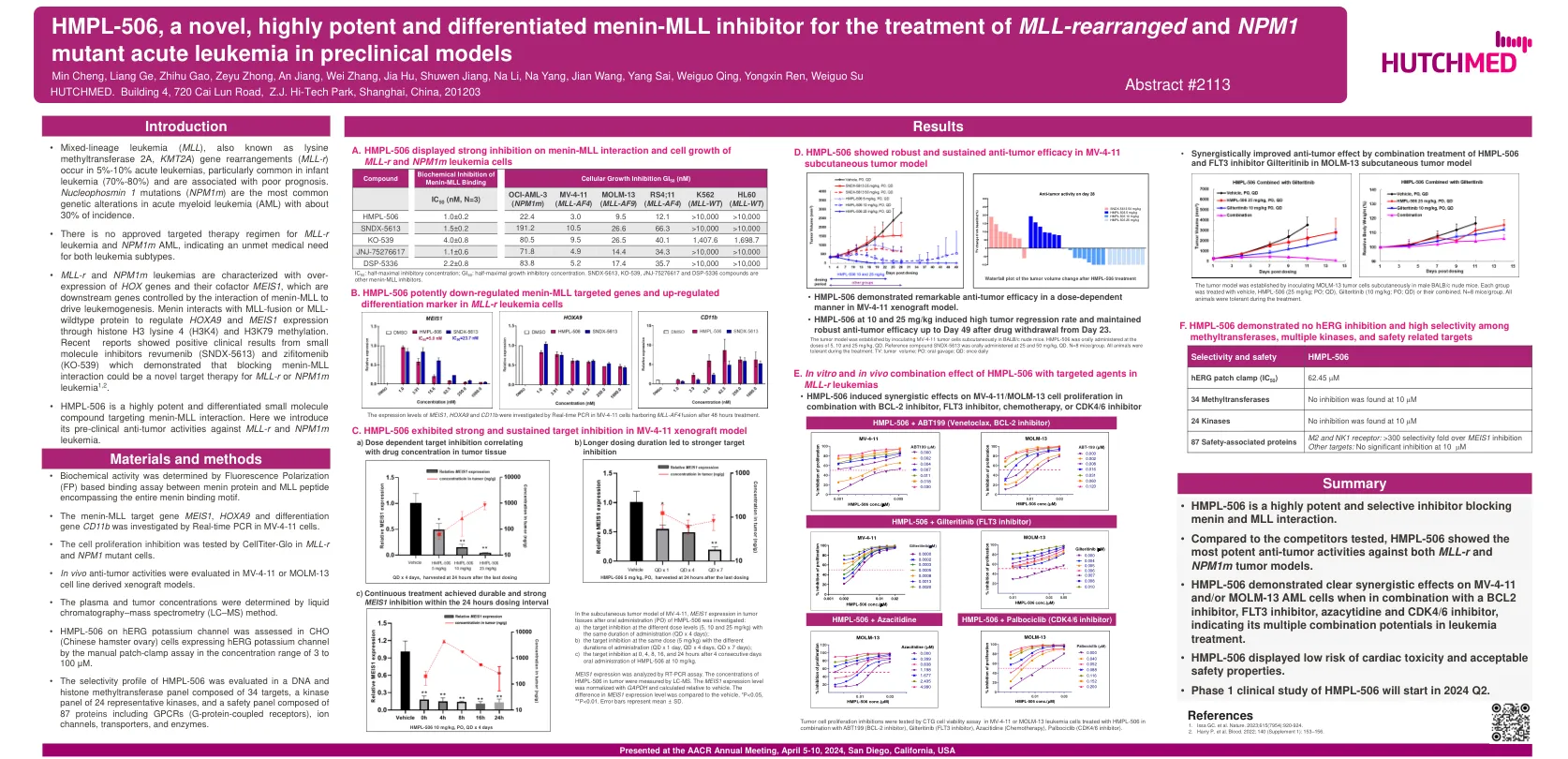
HMPL-506,一部小说,高度有效的和分化的Menin ...
HOX基因及其辅助因子Meis1的表达,它们是由Menin-Mll相互作用以驱动白血病发生的下游基因。Menin与MLL融合或MLL-野生型蛋白相互作用,通过组蛋白H3赖氨酸4(H3K4)和H3K79甲基化来调节HOXA9和MEIS1表达。最近的报道显示,小分子抑制剂rebumenib(SNDX-5613)和Zifitomenib(KO-539)的阳性临床结果,这些结果表明,阻断Menin-MLL相互作用可能是对MLL-R-R或NPM1M1M1M1M1M1M1M1M1M1,2的新型靶标治疗。

急性髓样白血病中的Smad1损失KMT2A
结果:在这里,我们报告了KMT2A :: AFF1和KMT2A :: MLLT3融合基因依赖性基因的下调Smad1(TGF-B信号轴转录因子)。SMAD1表达在大多数AML患者样品和包含两个融合基因KMT2A :: AFF1和KMT2A :: MLLT3的细胞系中丢失。SMAD1表达的丧失是通过将两个KMT2A融合基因引入造血干细胞和祖细胞中的。SMAD1的损失与具有KMT2A :: AFF1和KMT2A :: MLLT3的测试细胞中SMAD1启动子的H3K4me3水平显着降低。Smad1在具有KMT2A :: AFF1融合基因的细胞中的表达影响了细胞在体外的体外和影响kmt2a :: aff1细胞系MV4-11的体外植入。在MV4-11细胞中Smad1表达引起Hoxa9和Meis1的下调,这是通过TGF-B刺激加强的。 此外,在MV4-11细胞中,SMAD1的存在敏化细胞对TGF-B介导的G1暂停。在MV4-11细胞中Smad1表达引起Hoxa9和Meis1的下调,这是通过TGF-B刺激加强的。此外,在MV4-11细胞中,SMAD1的存在敏化细胞对TGF-B介导的G1暂停。

白血病洞察——2021 年 12 月
目前,急性白血病对menin的依赖模式与HOX基因及其辅因子MEIS1的过度表达有关;因此,这种基因表达谱可用作menin抑制反应的生物标志物。然而,鉴于目前缺乏经过验证的检测方法,以前显示具有这种基因表达特征的白血病基因型可用作反应的替代标志物。除了突变的NPM1和KMT2Ar外,AML中的其他基因型或复发性细胞遗传学异常也具有这种表达特征(表1)。举一个例子,涉及核孔复合体98(NUP98)的重排的白血病很少见,但预后不良,在临床前模型和患者样本中HOXA9过度表达,临床前数据显示对menin抑制易感。

优先考虑十一-十九种白血病抑制剂作为治疗急性髓系白血病的潜在候选药物
图 4. 72 小时 ENL 抑制剂处理后,(a) MOLM-13 (b) MV4-11 (c) Jurkat 和 (d) HEK293T 细胞的存活率。(e) 72 小时抑制剂 13 处理后,MOLM-13、MV4-11、Jurkat 和 HEK293T 细胞的细胞存活率比较。(f) MOLM-13 (g) MV4-11 和 (h) Jurkat 细胞在 10 µM ENL 抑制剂作用下的增殖。(i) 在指定温度下,用 10 μM (+) 或 DMSO (-) 中的 13 处理的 MOLM-13 (顶部) 和 MV4-11 (底部) 细胞中的 CETSA。β-肌动蛋白用作上样对照。用 13 或 DMSO 阴性对照处理的 (j) MOLM-13 和 (k) MV4-11 细胞中 HOXA9、MEIS1、MYB 和 MYC 基因表达的 qRT-PCR 分析。 *P < 0.05,**P < 0.01,***P < 0.001,****P < 0.0001。不显著 (ns) P > 0.05。

2024年梅宁抑制剂的更新。一类针对KMT2A重新培养和NPM1突变的急性髓样白血病
摘要:Menin抑制剂是目前正在临床开发中的新型和有前途的药物,其针对HOX/MEIS1转录程序,这对于在组蛋白赖氨酸N-甲基转移酶2A重新培训(KMT2AR)和NPM1-氧化(NPM1)氧化(NPM1M1M1M1MUT)尖锐leukemias中至关重要。这种新型药物的作用机理是基于MENIN – KMT2A复合物的破坏(由染色质重塑蛋白组成),从而导致表达KMT2A或突变NPM1的AML细胞的分化和凋亡。迄今为止,这种新的药物已在I阶段和II期临床试验中进行了测试,无论是单独的,并与协同药物结合使用,在经过预先治疗的急性白血病患者的缓解率和安全性方面,显示出令人鼓舞的结果。在这篇简短的综述中,我们总结了有关梅宁抑制剂的关键发现,重点介绍了有关急性髓细胞性白血病治疗的作用机理和初步的临床数据,尤其是这种有希望的新药物,尤其是Revumenib和Ziftomenib。