XiaoMi-AI文件搜索系统
World File Search SystemMorimoto

Morimoto Mitsuru教授的ZA:Suzuki takuji肺干细胞生物学和疾病
肺组织具有各种类型的上皮组织干细胞,在组织稳态中起着至关重要的作用,并因吸入化学颗粒以及病毒/细菌感染引起的急性损伤而再生。由于如此重要的作用,组织干细胞的功能障碍与呼吸道疾病有关。在今晚的研讨会上,我将介绍我们目前关于两个肺部干细胞的发现。气道基底细胞和牙槽II型(AT2)细胞。1)基底细胞通过从缓慢的循环转变为增殖,然后又回到缓慢的循环中,从而导致成人组织再生。尽管持续增殖会导致肿瘤发生,但调节这些转变的分子机制仍然未知。使用发育中的鼠气祖细胞的时间单细胞转录组学,我们发现TGF-β-ID2轴通常调节发育和再生过程中基础细胞中基础细胞中的增殖转变,并且其微调对正常再生至关重要,同时避免基础细胞增生。2)肺泡是肺纤维化起源的主要根源,已广泛研究了分子病因。调节肺泡上皮细胞纤维化状态的机制仍然难以捉摸。为了阐明上皮损伤和肌纤维细胞分化之间的因果关系,我们使用AT2干细胞培养建立了一个基于器官的肺纤维化模型。我们发现核心细胞系统在肺纤维发生中起着核心作用。该模型系统可用于研究较少炎症的肺纤维化的初始诱导,包括特发性肺纤维化。



第 18 届 NICT TDC 研讨会论文集(鹿岛,2020 年 10 月 1 日)
1987 年,无线电研究实验室(RRL,现 NICT)决定建造鹿岛 34 m 天线作为西太平洋干涉仪的主站。当时,日本国立天文台(NAOJ)的野边山射电天文台参与了使用野边山 45 m 的全球毫米波 VLBI,并刚刚开始 VLBI 观测。一套 Mark-3 记录器从野边山运到臼田站,使用臼田 64 m 进行了首次与 TDRS 卫星的空间 VLBI 实验,并成功进行了条纹检测。然而,在日本,独立的天文 VLBI 观测研究尚未实现。听到鹿岛34米天线建设的消息后,森本教授(图2)注意到鹿岛34米天线的面精度为170μ,对毫米波VLBI观测非常有效。森本教授向RRL提议与NAOJ合作进行毫米波VLBI研究,于是联合研究开始了。NAOJ决定利用RRL拥有的43GHz冷却接收机杜瓦瓶,制造出世界上第一台43GHz冷却HEMT接收机(图3),联合研究于1989年开始。KNIFE实验与34米天线的启动和测试同时开始。虽然

免疫・微生物学教室セミナー Evolutionary trajectory of ...
Ryo Morimoto,医学博士,博士莫里莫托(Morimoto)发展生物学系(BOEHM LAB),MAX PLANCK免疫生物学和表观遗传学研究所日期和时间:2024年1月24日,星期三,10:30-11:30地点:药学毕业生,医学院毕业生,从10:30:30:30:药房,来自四楼的药房,药学研究生院,语言:日语联系人:Hori Shohei(Ext。) 24820)摘要:针对病原体的人类自卫系统由两个主要的武器组成:先天和适应性免疫组织化学。 与先天抗原受体(AGR)识别模式识别的组合以及淋巴细胞表达的自适应AGR的预期模式对于建立我们精心策划的自卫以及保持体内平衡是必不可少的。 值得注意的是,这种复杂的系统仅在脊椎动物物种中建立,而大多数无脊椎动物物种仅取决于先天的免疫反应。 为了了解我们“现代”系统的进化紧急和轨迹,我想引入在无知的脊椎动物(hagfishes and Lampreys)中发现的替代自适应免疫学,这是一个与颚式脊椎动物(从鲨鱼到人类)5亿年前分离的小姐妹群体。 虽然它们的替代系统具有适应性免疫学的体液和细胞臂,但AGR的分子实体基于富含亮氨酸的重复模块。 与我们的V(d)J重组通过RAG1/2活性相反,AGRS的组装是通过基于胞苷脱氨酶(CDA)功能的一系列基因转换步骤来实现的。Ryo Morimoto,医学博士,博士莫里莫托(Morimoto)发展生物学系(BOEHM LAB),MAX PLANCK免疫生物学和表观遗传学研究所日期和时间:2024年1月24日,星期三,10:30-11:30地点:药学毕业生,医学院毕业生,从10:30:30:30:药房,来自四楼的药房,药学研究生院,语言:日语联系人:Hori Shohei(Ext。24820)摘要:针对病原体的人类自卫系统由两个主要的武器组成:先天和适应性免疫组织化学。与先天抗原受体(AGR)识别模式识别的组合以及淋巴细胞表达的自适应AGR的预期模式对于建立我们精心策划的自卫以及保持体内平衡是必不可少的。值得注意的是,这种复杂的系统仅在脊椎动物物种中建立,而大多数无脊椎动物物种仅取决于先天的免疫反应。为了了解我们“现代”系统的进化紧急和轨迹,我想引入在无知的脊椎动物(hagfishes and Lampreys)中发现的替代自适应免疫学,这是一个与颚式脊椎动物(从鲨鱼到人类)5亿年前分离的小姐妹群体。虽然它们的替代系统具有适应性免疫学的体液和细胞臂,但AGR的分子实体基于富含亮氨酸的重复模块。与我们的V(d)J重组通过RAG1/2活性相反,AGRS的组装是通过基于胞苷脱氨酶(CDA)功能的一系列基因转换步骤来实现的。
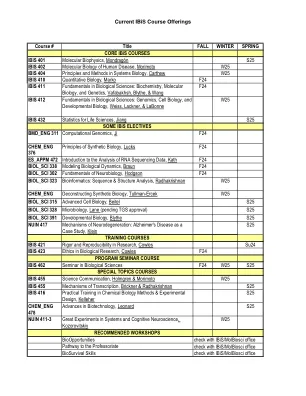
当前的IBIS课程产品
TRAINING COURSES IBIS 421 Rigor and Reproducibility in Research, Cowles Su24 IBIS 423 Ethics in Biological Research, Cowles F24 PROGRAM SEMINAR COURSE IBIS 462 Seminar in Biological Sciences F24 W25 S25 SPECIAL TOPICS COURSES IBIS 455 Science Communication, Holmgren & Morimoto W25 IBIS 455 Mechanisms of Transcription, Brickner &Radhakrishnan S25 IBIS 416化学生物学方法与实验设计的实践培训,Kelleher

Konezumi,Dajin,帮助创建高质量基因组编辑的小鼠...
Kuno,A.,Ikeda,Y.,Ayabe,S.,Kato,K.,Sakamoto,K.,M.,M.,A. div>dajin使多重基因分型同时验证预期和意外的目标基因组编辑结果。 div>PLOS Biology,20(1),E3001507。 div>
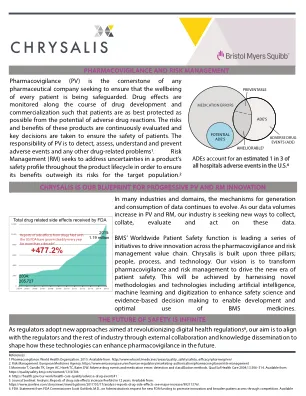
chrysalis factsheet
参考文献:1个药物宣传。世界卫生组织。2015。可从:http://www.who.int/medicines/areas/quality_safety/safety_e-ceracy/pharmvigi/en/2。风险管理。欧洲药品局。https://www.ema.europa.eu/en/human-regulatory/marketing-authorisation/pharmacovigilance/risk-management 3 Morimoto T,Gandhi TK,Seger tk,Seger AC,AC,HSIEH TC,BATES DC,BATES DW。不良药物事件和药物错误:检测和分类方法。Qual Saf Health Care 2004; 13:306–314。可从:https://qualitysafety.bmj.com/content/13/4/306 4. https://health.gov/our-work/work/work/health-care-quare-quality/ardverse-drug-events#1 5。日记哨兵。分析:药物副作用的报道在12年内增加了五个。可从:https://www.jsonline.com/story/news/investigations/2017/03/17/analsiss-reports-reports-drug-side-eects-see-see-major-see-major-increase/99211376/。6。fda。,FDA专员Scott Gottlieb,M.D。的声明,就管理要求提供新的FDA资金,以促进创新并通过竞争扩大患者访问权限。可用

关于研究主题哺乳动物精子发生的社论:遗传和环境因素
精子发生是一个复杂且严格调节的过程,其中包括精子的增殖,精子分化为精子细胞,生产精子的减数分裂分裂,圆形精子成熟,精子的成熟以及高度专业的成熟精子的精子释放以及释放。这些事件中的任何一个异常都可能导致影响生育能力的精子发生障碍。精子发生障碍可能是由遗传和非遗传因素引起的,其中遗传因素占15%至30%,非遗传学占70% - 85%(O'Flynn O'Brien等,2010; Neto等,2016)。值得注意的是,作为非遗传学的环境因素对于精子发生很重要,因为男性生殖系统,尤其是精子发生似乎对环境危害特别敏感(Vecoli等,2016)。本研究主题包括七个原始文章和一项迷你审查,以增强和扩展我们对这些因素和机制的了解。精子干细胞(SSC)是最原始的生殖细胞,通过自我更新和连续分化为精子细胞,在睾丸中产生精子(Kubota and Brinster,2018),它们通过自我更新和连续分化来维持精子发生。Wu等人的研究。发现GPX3调节人类SSC的增殖和凋亡。作者表明,GPX3在人类SSC中高度表达,其敲低抑制了细胞增殖。此外,GPX3与CXCL10相互作用,并且它们的敲低表型在人类SSC系列中是一致的。结果表明GPX3和CXCL10对于SSC自我更新至关重要。有一些关于外部环境因素对SSC自我更新和分化的影响的研究。先前的研究表明,缺氧对SSC的增殖有益(Morimoto等,2021)。在此研究主题中,Gille等人。研究了缺氧如何影响SSC的增殖和分化。作者证明,当O2张力≤1%时,SSC显示出轻微的分化偏置和增殖的减少,这与Morimoto等人的结果一致。(2021)。减数分裂过程中发生了几个重要事件,包括DNA复制,染色质冷凝,DSB形成和DSB修复。这些事件不是减数分裂的独家,并且发生在体细胞周期中,并且已证明核肌动蛋白与这些事件有关。但是,没有研究来阐明核肌动蛋白和减数分裂之间的关系。在此研究主题中,Petrusová等。提供了一个迷你审查,以阐明核肌动蛋白在预言I

《AI哲学图谱》【综合讨论/下篇】人工智能——哲学对应图谱
[查尔默斯 01] 大卫·查尔默斯,Hajime Hayashi 译:《意识:寻找大脑和精神的基本理论》,白洋社(2001) [克拉克 22] 安迪·克拉克,Takashi Ikegami 和 Gentaro Morimoto 译:《显现的存在:大脑、身体和世界的重新整合》,Hayakawa Publishing(2022) [笛卡尔 67] 勒内·笛卡尔,Taro Ochiai 译:《方法论》,Iwanami Bunko(1967) [德勒兹 12] 吉尔斯·德勒兹和菲利克斯·瓜塔里,Osamu Zaitsu 译:《什么是哲学?》,Kawade Bunko(2012) [丹尼特 96] 丹尼尔·丹尼特,Tadashi Wakashima 和 Manabu Kawata 译:《意图》 “态度”的哲学——人能读懂别人的行为吗? ,白洋社(1996) [Ganassia 19] Ganassia Jean-Gabriel,伊藤直子译:埋葬虚假的AI神话“奇点”,早川出版(2019) [Heidegger 13] Heidegger Martin,熊野澄彦译:存在与时间,岩波文库(2013) [Hume 04] 休谟·戴维,斋藤繁雄、一之濑正树译:人类智力研究——附人性论概要,法政大学出版会(2004) [Husserl 79] 胡塞尔·埃德蒙,渡边次郎译:理念 I-I 纯粹现象学概论,美铃书房(1979) [ Husserl 01] 埃德蒙德·胡塞尔,滨涡达二译:《笛卡尔的沉思》,岩波文库(2001) [Jung 16] 卡尔·荣格,林道吉译:《个体化与曼荼罗(新版)》,美铃书房(2016) [Kant 60] 伊曼纽尔·康德,篠田秀夫译:《纯粹理性批判》,岩波文库(1960) [Kurzweil 07] Ley Kurzweil,井上健、小野木章监修翻译