XiaoMi-AI文件搜索系统
World File Search SystemNRF2

研究亨廷顿疾病管理的整体自然策略
摘要。亨廷顿氏病(HD)是一种多方面的神经系统疾病,呈现出复杂的临床情况。一种称为亨廷顿疾病的常染色体显性神经退行性疾病是由CAG数量增加(细胞质 - 腺苷 - 瓜氨酸)重复序列引起的,这会导致突变的亨廷顿蛋白(MHTT)产生人类生物的神经元死亡和心理障碍。结束体征和症状可能包括重大体重减轻,吞咽困难或呼吸困难,复发性抽吸肺炎,健康状况下降和不受控制的疼痛。在包括HD在内的大多数神经系统疾病中,神经组织中ROS过多的ROS(活性氧)被认为是重要的危险因素。转录调控,免疫系统和线粒体功能都被MHTT破坏了。尽管天然产品在改善症状方面已经有希望,但重要的是要注意,尚无单一的“植物性遗产”与其治疗性干预有明确的联系。然而,某些天然发生的化合物在临床前研究中表现出了有希望的结果。本文重点介绍了一些通过广泛的生物学活性具有多种神经保护作用的植物成分。通过刺激NRF2(核因子2相关因子)途径,并抑制NF-κB(核因子Kappa-Light-chain-chain-Enhancer),astaxanthin,berberine和sulfarophane和磺烷会增加抗氧化剂和抗炎性活性,并产生NeuroRrotsection。姜黄素会导致金属螯合作用和活性氧的下降,这无疑是阻碍和管理引起神经退行性疾病(包括HD)的疾病的重要过程之一。这会影响HSP(热休克蛋白)的上调,这有助于HD管理。纳林蛋白通过自由基清除来降低氧化应激水平和炎症水平,NF-κB刺激细胞存活,并通过上调抗凋亡基因的表达和下调凋亡基因来防止凋亡。
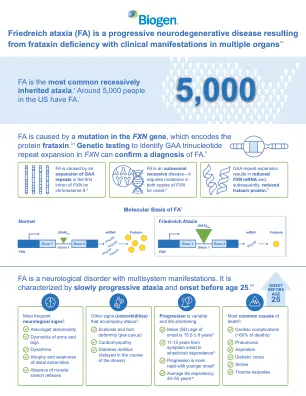
弗里德赖希共济失调 (FA) 是一种由 frataxin 缺乏引起的进行性神经退行性疾病,临床表现涉及多个器官 1
ARE,抗氧化反应元件;ATP,三磷酸腺苷;DNA,脱氧核糖核酸;FA,弗里德赖希共济失调;GAA,鸟嘌呤腺嘌呤腺嘌呤;ISC,铁硫簇;Keap1,Kelch 样 ECH 相关蛋白 1;Nrf2,核因子红细胞 2 相关因子 2;OXPHOS,氧化磷酸化;ROS,活性氧;SD,标准差。参考文献:1. 弗里德赖希共济失调研究联盟。什么是 FA?可从 https://www.curefa.org/understanding-fa/what-isfriedreichs-ataxia/ 获取。访问日期:2024 年 11 月。2. Koeppen AH。J Neurol Sci。2011;303(1-2):1-12。3. Campuzano V 等人。Hum Mol Genet。 1997;6(11):1771-1180。 4.Nachun D 等人。哈姆·摩尔·热内特。 2018;27(17):2965-2977。 5.弗里德赖希共济失调研究联盟。 Friedreich 共济失调临床管理指南 (FRDA)。可从 https://frdaguidelines.org/ 获取。访问时间:2024 年 11 月。 6. Campuzano V 等人。科学 。 1996;271(5254):1423-1427。 7.Gatchel JR 等人。纳特·热内特。 2005;6(10):743-755。 8. Bürk K. 小脑共济失调。 2017;4:4。 9.潘道夫·M·尼罗尔·吉内特。 2020;6(3):e415。 10. 汉森 E 等人。世界心脏病杂志。 2019;11(1):1-12。 11.Chiang S 等。神经化学国际公司。 2018;117:35-48。 12. González-Cabo P,帕劳 F. J Neurochem。 2013;126(补编1):53-64。 13. Llorens JV 等。神经科学前沿。 2019;13:75。 14. Petrillo S 等人。国际分子科学杂志。 2017;18(10):2173。 15.D'Oria V 等人。国际分子科学杂志。 2013;14(4):7853–7865。 16. Itoh K 等人,基因发育. 1999;13(1):76-86。17. Santos R 等人,抗氧化还原信号. 2010;13(5):651-690。

海报展示(截至 2023 年 9 月 14 日)
海报展示(截至 2023 年 9 月 14 日)海报会议 B 10 月 13 日星期五 | 下午 12:30-下午 4:00 第 2 层,展览厅 D B002:FHD-286 在 AML 或 MDS 患者中开展的 1 期研究中的药效学和抗肿瘤机制。Mike Collins,Foghorn Therapeutics,美国马萨诸塞州剑桥。B003:从晚期癌症患者的肿瘤组织和 Tempus 基因组数据库的液体活检中收集的 TSC1 和/或 TSC2 变异的真实世界 (RW) 表征和频率。David J. Kwiatkowski,布莱根妇女医院,美国马萨诸塞州波士顿。B004:NF-κB 和 NRF2 信号之间的分子串扰影响 HPV 相关头颈癌的预后。Aditi Kothari,北卡罗来纳大学,美国北卡罗来纳州教堂山。 B005:分子分析和 ESCAT 分类对患者结果的影响:居里研究所分子肿瘤委员会的经验。Maud Kamal,法国巴黎居里研究所。B006:通过邻近连接试验评估的高 RAS-RAF 结合与 NSCLC 对 KRAS G12C 抑制剂的敏感性有关。Ryoji Kato,美国佛罗里达州坦帕市 H. Lee Moffitt 癌症中心和研究所。B007:NCI-MATCH 试验 (EAY131) 中肿瘤组织和血浆基因分型之间的一致性。Mohamed A. Gouda,德克萨斯大学 MD 安德森癌症中心,美国德克萨斯州休斯顿。B008:机器学习支持对具有光谱重叠的共定位多重 IHC 信号进行量化。Waleed Tahir,PathAI,美国马萨诸塞州波士顿。 B009:基于面板的同源重组缺陷突变特征与转移性去势抵抗性前列腺癌对 PARP 抑制的反应有关。Daniel Boiarsky,塔夫茨医学中心,美国马萨诸塞州波士顿。B010:使用加性多实例学习模型对 H&E 全幻灯片图像中的基因表达特征进行空间分辨预测。Chintan Parmar,PathAI,美国马萨诸塞州波士顿。B011:GDF-15 是上皮样血管内皮瘤侵袭性的生物标志物,并通过 ATF4 抑制被雷帕霉素下调。Alessia Beretta,意大利米兰国家肿瘤研究所 IRCCS 基金会。 B012:验证 OncoSignature 检测,这是一种针对 ACR-368 的响应预测定量多重免疫荧光检测,用于预测癌症患者对 CHK1/2 抑制剂 ACR-368 的敏感性。Michail Shipitsin,Acrivon Therapeutics,美国马萨诸塞州沃特敦。B013:乳腺癌 Notch 转录组特征的鉴定。Felix Geist,默克集团医疗保健业务,德国达姆施塔特。B014:非小细胞肺癌患者 Nectin-4 蛋白表达的特征。Sean Santos,Bicycle Therapeutics,美国马萨诸塞州剑桥。

2024 年静修海报任务
基于网络的表观遗传和转录组景观整合揭示了人类 T 滤泡辅助细胞分化背后的分子程序 62. Prabal Chhibbar、Priyamvada Guha Roy、Munesh K. Harioudh、Daniel J. McGrail、Donghui Yang、Harinder Singh、Reinhard Heinterleitner、Yi-Nan Gong、S. Stephen Yi、Nidhi Sahni、Saumendra N. Sarkar 和 Jishnu Das 使用结构解析的蛋白质网络揭示 COVID-19 中细胞类型特异性免疫调节变体和相关分子表型 63. Andrew W. Liu、Youran Zhang、Chien-Sin Chen、Sumeyye Ozyaman、Tara N. Edwards、Torben Ramcke、Eric S. Weiss、Jacob E. Gillis、Colin R. Laughlin、Catherine M. Phelps、Simran K. Randhawa、Marlies Meisel、Tina L. Sumpter 和 Daniel H. Kaplan 搔痒会加剧过敏性炎症并通过神经源性肥大细胞活化增强宿主防御 64. Dan Xue、Mengqi Huang、Yuechen Zhou、Eleanor Valenzi 和 Robert Lafyatis 揭示系统性硬化症中 TGFB3 上调的原因:从染色质可及性和转录因子分析中获得的见解 65. Jane C. Siwek、Alisa A. Omelchenko、Prabal Chhibbar、Sanya Arshad、Iliyan Nazarali、Kiran Nazarali、AnnaElaine Rosengart、Javad Rahimikollu、Jeremy Tilstra、Mark J. Shlomchik、David R. Koes、Alok V. Joglekar 和 Jishnu Das 滑动窗口相互作用语法(SWING):一种用于肽和蛋白质相互作用的广义相互作用语言模型 66. Danica Lee、Urekha Karri、Prabal Chhibbar、Priyamvada Guha Roy、Jishnu Das 和Daniella Schwartz 与 A20 单倍体不足(HA20)相关的 TNFAIP3 变异的基因型到表型分析表明患病率高于预期 67. Jacob E. Gillis、Chien-Sin Chen、Caitlin O. Bacon、Tara Edwards、Andrew W. Liu、Eric S. Weiss、Jonathan A. Cohen 和 Daniel H. Kaplan CGRP 在神经源性皮肤炎症中的作用 68. Hanxi Xiao、Niranjana Natarajan、Partha Dutta 和 Jishnu Das 揭示心肌梗死中免疫细胞的空间微环境动态 69. Anthony J. Bragoli、Zhangguo Chen、Karen Siddoway 和 Jing H. Wang Ly6C 和 Ly6A 在化疗治疗的癌细胞诱导的 CD8 + T 细胞激活中的作用(不依赖于 MHC I 类) 70. Amanda Lee、Surya P. Pandey、Colin R. Laughlin、Alex McPherson 和 Marlies Meisel 定义 AhR 和 Nrf2 在抗肿瘤免疫的共生免疫调节中的作用

SGLT2抑制剂的心脏保护和抗癌作用
Akt¼蛋白激酶B; ALP¼碱性磷酸酶; a-sma¼a -smooth肌肉肌动蛋白; AMPK¼腺苷单磷酸 - 活化的蛋白激酶; ANP¼14钠肽; Arn¼血管紧张素受体Neprilysin抑制剂; AST¼天冬氨酸氨基转移酶; ATF-4¼激活转录因子4; BAX¼Bcl-2相关X蛋白; B-MHC¼B-肌球蛋白重链; bohb¼b-羟基丁酸酯; BNP¼B型纳特里尿肽; CAT¼过氧化氢酶; CFR¼冠状动脉储备; CK-MB¼肌酸激酶MB; CRS¼心脏综合征; CTNT¼心脏肌钙蛋白T;潮湿¼损伤相关的分子模式; dox¼阿霉素; ECG¼心电图; ef¼射血分数; EIF-2a¼真核生物起始因子2 a; Er¼内质网; ERK¼1.1.1/1/14; FGF¼FIMBLAST生长因子; FS¼部分缩短; g-csf¼1/1/14 GM-CSF¼1/1/1/14 GRP78¼葡萄糖调节的蛋白78; HTN¼高血压; I.P.¼腹膜内; IL¼白痴; IL¼白痴; IL¼白痴; iNOS¼诱导一氧化氮合酶; LDH¼14乳酸脱氢酶; LV¼左心室; lvedd¼左心室末端直径; lvesd¼左心室末端音直径; LVIDD¼左心内直径在末端末端;末端收缩处的LVIDS¼左心内直径; MDA¼MALONDIALLEDEDEDE; MMP¼基质金属肽酶; MPO¼髓过氧化物酶;雷帕霉素的mtor¼哺乳动物靶标; mybpc3¼结合蛋白C3; MyD88¼髓样差异反应88; NCD¼正常食物饮食; NF-kb¼核因子kappa-b; NLRP3¼NOD样受体蛋白3;无¼一氧化氮; NOX-1¼NADPH氧化酶1; NOX-2¼NADPH氧化酶2; NRF2¼核因子红细胞2 - 相关因子2; NT-Proanp¼n末端Pro - 心房纳地肽; NT-PROBNP¼N末端Pro - B型纳地尿肽; p38¼p38有丝分裂原激活的蛋白激酶; PARP¼聚(二磷酸腺苷 - 核糖)聚合酶; PERK¼蛋白激酶R样性内质网激酶; PGC¼过氧化物酶体增殖物 - 激活的受体共激活剂; PI3K¼磷酸肌醇3-激酶; PPAR¼过氧化物酶体增殖物 - 活化受体; QTC¼校正的QT; SIRT1¼SIRTUIN1; Sirt3¼Sirtuin3; Smad3¼母亲反对脱皮的同源物3; SOD¼超氧化物歧化酶; TGF¼转化生长因子; TLR9¼Toll样受体9; TNF¼肿瘤坏死因子; XO¼黄嘌呤氧化酶;其他缩写如表1所示。

海报展示(截至 9/19/23)
海报展示(截至 2023 年 9 月 19 日)海报会议 B 星期五,10 月 13 日 | 下午 12:30-下午 4:00 第 2 层,展览厅 D B002:FHD-286 在 AML 或 MDS 患者中开展的 1 期研究中的药效学和抗肿瘤机制。Mike Collins,Foghorn Therapeutics,美国马萨诸塞州剑桥。B003:从晚期癌症患者的肿瘤组织和 Tempus 基因组数据库的液体活检中收集的 TSC1 和/或 TSC2 变异的真实世界 (RW) 表征和频率。David J. Kwiatkowski,布莱根妇女医院,美国马萨诸塞州波士顿。B004:NF-κB 和 NRF2 信号之间的分子串扰影响 HPV 相关头颈癌的预后。Aditi Kothari,北卡罗来纳大学,美国北卡罗来纳州教堂山。 B005:分子分析和 ESCAT 分类对患者结果的影响:居里研究所分子肿瘤委员会的经验。Maud Kamal,法国巴黎居里研究所。B006:通过邻近连接试验评估的高 RAS-RAF 结合与 NSCLC 对 KRAS G12C 抑制剂的敏感性有关。Ryoji Kato,美国佛罗里达州坦帕市 H. Lee Moffitt 癌症中心和研究所。B007:NCI-MATCH 试验 (EAY131) 中肿瘤组织和血浆基因分型之间的一致性。Mohamed A. Gouda,德克萨斯大学 MD 安德森癌症中心,美国德克萨斯州休斯顿。B008:机器学习支持对具有光谱重叠的共定位多重 IHC 信号进行量化。Waleed Tahir,PathAI,美国马萨诸塞州波士顿。 B009:基于面板的同源重组缺陷突变特征与转移性去势抵抗性前列腺癌对 PARP 抑制的反应有关。Daniel Boiarsky,塔夫茨医学中心,美国马萨诸塞州波士顿。B010:使用加性多实例学习模型对 H&E 全幻灯片图像中的基因表达特征进行空间分辨预测。Chintan Parmar,PathAI,美国马萨诸塞州波士顿。B011:GDF-15 是上皮样血管内皮瘤侵袭性的生物标志物,并通过 ATF4 抑制被雷帕霉素下调。Alessia Beretta,意大利米兰国家肿瘤研究所 IRCCS 基金会。 B012:验证 OncoSignature 检测,这是一种针对 ACR-368 的响应预测定量多重免疫荧光检测,用于预测癌症患者对 CHK1/2 抑制剂 ACR-368 的敏感性。Michail Shipitsin,Acrivon Therapeutics,美国马萨诸塞州沃特敦。B013:乳腺癌 Notch 转录组特征的鉴定。Felix Geist,默克集团医疗保健业务,德国达姆施塔特。B014:非小细胞肺癌患者 Nectin-4 蛋白表达的特征。Sean Santos,Bicycle Therapeutics,美国马萨诸塞州剑桥。

海报演示(截至9/14/23)
海报演示文稿(截至9/14/23)10月13日(星期五)海报会议B | 12:30 pm-4:00 PM 2级,展览馆D B002:在AML或MDS受试者中,在1期研究中,FHD-286的药效学和抗肿瘤机制。Mike Collins,Foghorn Therapeutics,美国剑桥,美国。b003:从肿瘤组织和液体活检中收集的TSC1和/或TSC2改变的现实世界(RW)表征和频率是晚期癌症患者的TEMPUS基因组数据库的液体活检。美国马萨诸塞州波士顿的杨百翰和妇女医院的戴维·J·Kwiatkowski。b004:NF-κB和NRF2信号之间的分子串扰会影响与HPV相关的头颈癌的预后。Aditi Kothari,UNC,美国北卡罗来纳州教堂山。b005:分子分析和ESCAT分类对患者结局的影响:库里学院分子肿瘤板的经验。莫德·卡马尔(Maud Kamal),法国巴黎库里学院。b006:通过接近连接测定评估的高RAS-RAF结合与NSCLC中对KRAS G12C抑制剂的敏感性有关。Ryoji Kato,H。LeeMoffitt癌症中心和研究所,佛罗里达州坦帕。 B007:NCI匹配试验中肿瘤组织与血浆基因分型之间的一致性(EAY131)。 Mohamed A. Gouda,德克萨斯大学医学博士Anderson癌症中心,美国德克萨斯州,美国。 b008:机器学习启用了具有光谱重叠的共定位多重IHC信号的量化。 Waleed Tahir,Pathai,波士顿,美国马萨诸塞州。 Daniel Boiarsky,塔夫茨医疗中心,美国马萨诸塞州。 Chintan Parmar,Pathai,波士顿,美国马萨诸塞州。Ryoji Kato,H。LeeMoffitt癌症中心和研究所,佛罗里达州坦帕。B007:NCI匹配试验中肿瘤组织与血浆基因分型之间的一致性(EAY131)。Mohamed A. Gouda,德克萨斯大学医学博士Anderson癌症中心,美国德克萨斯州,美国。 b008:机器学习启用了具有光谱重叠的共定位多重IHC信号的量化。 Waleed Tahir,Pathai,波士顿,美国马萨诸塞州。 Daniel Boiarsky,塔夫茨医疗中心,美国马萨诸塞州。 Chintan Parmar,Pathai,波士顿,美国马萨诸塞州。Mohamed A. Gouda,德克萨斯大学医学博士Anderson癌症中心,美国德克萨斯州,美国。b008:机器学习启用了具有光谱重叠的共定位多重IHC信号的量化。Waleed Tahir,Pathai,波士顿,美国马萨诸塞州。Daniel Boiarsky,塔夫茨医疗中心,美国马萨诸塞州。 Chintan Parmar,Pathai,波士顿,美国马萨诸塞州。Daniel Boiarsky,塔夫茨医疗中心,美国马萨诸塞州。Chintan Parmar,Pathai,波士顿,美国马萨诸塞州。b009:基于面板的同源重组缺乏症的基于面板的突变特征,并响应转移性cast割的前列腺癌对PARP抑制作用。b010:使用添加性多个实例学习模型在H&E H&E整个幻灯片图像中基因表达特征的空间分辨预测。b011:GDF-15是上皮性血管内皮瘤侵略性的生物标志物,并由Sirolimus通过ATF4抑制而受到调节。Alessia Beretta,Fondazione Irccs Istituto Nazionale dei tumori,意大利米兰。b012:验证Oncosignature Assay是一种ACR-368级的反应预测定量多重多重免疫荧光测定法,以预测对CHK1/2抑制剂ACR-368的敏感性。Michail Shipitsin,Acrivon Therapeutics,美国沃特敦。B013:鉴定乳腺癌的凹痕转录组学特征。Felix Geist,德国达姆施塔特默克KGAA的医疗保健业务。b014:非小细胞肺癌患者中蜜素-4蛋白表达的表征。Sean Santos,自行车治疗学,美国剑桥。

A 部分。个人信息 名字 Rosa Maria ...
1. Erana-Perez Z、Igartua M、Santos-Vizcaino E*、Hernandez RM* (AC) 。差异蛋白质和 mRNA 货物装载到工程化大细胞外囊泡和小细胞外囊泡中揭示了体外和体内试验中的差异。J Control Release 379: 951 (2025) 影响因子:11.467,Q1。2. Las Heras K、Garcia-Orue I、Aguirre JJ、de la Caba K、Guerrero P、Igartua、Edorta Santos-Vizcaino M*、Hernandez RM* (AC) 。载有来自毛囊或脂肪组织的人类间充质基质细胞的大豆蛋白/β-几丁质海绵状支架可促进糖尿病慢性伤口愈合。Biomater Adv 155: 213682 (2023)。影响因子:7.9,第一季度。3. Las Heras K、Royo F、Garcia-Villacrosa C、Igartua M、Santos-Vizcaino、Falcon-Perez JM*、Hernandez RM* (AC)。毛囊来源的间充质基质细胞的细胞外囊泡:分离、表征和治疗慢性伤口愈合的潜力。干细胞研究与治疗 13:147 (2022)。影响因子:5.985,第一季度。4. Gonzalez-Pujana A、Vining KH、Zhang DKY、Santos-Vizcaino E、Igartua M、Hernandez RM (AC)、Mooney DJ (AC)。多功能仿生水凝胶系统可增强间充质基质细胞的免疫调节潜力。生物材料。257:120266 (2020)。如果:10.307,Q1。 5. 拉斯赫拉斯 K、桑托斯-比斯卡诺 E、加里多 T、古铁雷斯 FJ、阿吉雷 JJ、德拉卡巴 K、格雷罗 P、伊加图亚 M、埃尔南德斯 RM(AC)。大豆蛋白和甲壳质海绵状支架:从天然副产品到生物医学应用的细胞输送系统。绿色化学,22:3445-3460(2020)。如果:10.182,Q1。 6. 冈萨雷斯-普亚纳 A、桑托斯-维兹卡伊诺 E、加西亚-埃尔南多 M、埃尔纳兹-埃斯特拉达 B、M. 德潘科博 M、贝尼托-洛佩斯 F、伊加图亚 M、巴萨贝-德斯蒙特 L (AC)、埃尔南德斯 RM (AC)。基于细胞外基质蛋白微阵列的单细胞分辨率生物传感器:整合素分析和细胞-生物材料相互作用的表征。传感器和执行器,B:化学。299:126954 (2019)。影响因子:7.460,第一季度。7. Hernando S、Requejo C、Herran E、Ruiz-Ortega JA、Morera-Herreras T、Lafuente JV、Ugedo L、Gainza E、Pedraz JL、Igartua M (AC)、Hernandez RM (AC)。n-3 多不饱和脂肪酸在帕金森病部分病变模型中的有益作用:神经胶质细胞和 NRf2 调节的作用。神经生物学疾病 121:252-262 (2019)。影响因子:5.332,第一季度。 8. Garcia-Orue I、Santos-Vizcaino E、Etxabide A、Uranga j、Bayat A (AC).、Guerrero P、Igartua M、de la Caba K、Hernandez RM (AC)。用于伤口愈合的仿生明胶和明胶/壳聚糖双层水膜的开发。药剂学。 11(7):314-332(2019)。如果:4.699,Q1。 9. Hernando, S.、Herran, E.、Figueiro-Silva, J.、Pedraz JL、Igartua M.、Carro, E.,

胰腺β单元格:社论
胰腺β细胞通过作为胰岛素的主要来源来维持葡萄糖稳态方面起着关键作用。这些细胞负责胰岛素的合成,储存和释放,该胰岛素的合成,储存和释放是由于身体代谢状态的变化而受到严格调节。由于β细胞在糖尿病病理生理学中的核心作用,其生物学引起了科学界的显着兴趣。更好地了解β细胞生物学的多个方面可能会导致制定新的预防策略和治疗,从而延迟或停止疾病进展。这个特刊“胰腺β细胞”介绍了14篇文章的集合,其中包括五篇原始论文和9篇评论,突出了Beta细胞研究的各个方面。此问题的重点是控制β细胞质量膨胀和存活的分子机制,尤其强调了成熟β细胞功能的显着途径。贡献涵盖了广泛的主题,包括氧化应激对β细胞的影响[1-3],组织间通信[4-7],以及β细胞质量和功能的主调节剂[8-10]等。Mukai及其同事[1]回顾了氧化应激和β细胞抗氧化剂机制的作用,总结了β细胞中抗氧化酶的低表达和氧化应激如何损害胰岛素的分泌。作者建议核因子2与2相关因子2(NRF2)是β细胞抗氧化剂反应的主要调节剂。作者建议TMAO可能在糖尿病生成条件下介导一种补偿性的蛋白质作用。Wu及其同事[3]探索了在氧化应激条件下4-辛基伊替酸(4-OI)对胰腺β细胞的影响。研究人员发现,4-OI治疗减少了活性氧的产生,抑制细胞死亡途径激活和炎性细胞因子分泌,并逆转了缺氧诱导的细胞死亡,这表明4-OI可能在氧化应激条件下增强β细胞存活。此外,此问题还包括有关分子和其他组织中产生因素对β细胞功能的影响的文章。Krueger及其同事[4]研究了肠道微生物代谢产物三甲胺N-氧化物(TMAO)对功能性β细胞质量的影响。研究人员发现,尽管据报道患者的2型糖尿病(T2D)水平升高,但TMAO保护了β细胞功能并改善氧化和内质网应激。Fernandez-Millan及其同事[5]讨论了诸如T2D等代谢疾病病理学中的组织间交流的重要性,强调了如何理解Beta细胞与代谢和非代谢组织进行通信的方式提供了新的研究领域。他们强调了来自各种器官和组织对β细胞生物学的分泌因素的影响,这表明血时间交流可以为糖尿病研究提供新的机会。内分泌胰岛与胰腺中外分泌细胞的物理接近允许这些相邻细胞类型之间的旁分泌相互作用。作者强调了在这些情况下对糖尿病进行早期诊断的重要性。外分泌疾病对β细胞的影响是Ciochina and Floeagues的评论[6],它描述了慢性胰腺炎,急性胰腺炎,囊肿性肿瘤,胰腺癌,胰腺癌,胰腺癌,胰腺切除术以及Autoimmmune Pancreatiation and Autoimmune Pancreatiation如何影响Beta Celle和Diabect。Kryvalap及其同事[7]回顾了在外分泌胰腺中表达的蛋白酶和Serpin蛋白酶抑制剂对胰岛病理生理学的影响。作者探讨了对抑制或增强蛋白酶的反对意见

CRISPR/Cas9基因编辑技术在阿尔茨海默病研究中的应用
[4] Kisilevsky R. 从关节炎到阿尔茨海默病:关于淀粉样变性发病机制的最新概念。Can J Physiol Pharmacol,1987,65:1805-15 [5] György B、Lööv C、Zaborowski MP 等人。CRISPR/Cas9 介导的瑞典 APP 等位基因破坏作为早发性阿尔茨海默病的治疗方法。Mol Ther Nucleic Acids,2018,11:429-40 [6] Zetterberg H、Mattsson N. 了解散发性阿尔茨海默病的病因。Expert Rev Neurother,2014,14:621-30 [7] Jack CR Jr、Knopman DS、Jagust WJ 等人。阿尔茨海默病病理级联动态生物标志物的假设模型。Lancet Neurol,2010,9:119-28 [8] Ittner LM、Ke YD、Delerue F 等。tau 的树突状功能介导阿尔茨海默病小鼠模型中的淀粉样蛋白 β 毒性。Cell,2010,142:387-97 [9] Muralidar S、Ambi SV、Sekaran S 等。tau 蛋白在阿尔茨海默病中的作用:主要的病理因素。Int J Biol Macromol,2020,163:1599-617 [10] Wang X、Wang W、Li L 等。阿尔茨海默病中的氧化应激和线粒体功能障碍。 Biochim Biophys Acta, 2014, 1842: 1240-7 [11] Grothe M, Heinsen H, Teipel SJ. 成年年龄范围内以及阿尔茨海默病早期阶段胆碱能基底前脑萎缩。Biol Psychiatry, 2012, 71: 805-13 [12] He Y, Ruganzu JB, Jin H, et al. LRP1 敲低通过调节 TLR4/NF- κB/MAPKs 信号通路加重 Aβ 1-42 刺激的小胶质细胞和星形胶质细胞神经炎症反应。Exp Cell Res, 2020, 394: 112166 [13] Huang HC, Hong L, Chang P, et al.壳寡糖减弱Cu 2+诱导的细胞氧化损伤和细胞凋亡,涉及Nrf2激活。Neurotox Res,2015,27:411-20 [14] Tomljenovic L. 铝和阿尔茨海默病:经过一个世纪的争论,是否存在合理的联系?J Alzheimers Dis,2011,23:567-98 [15] Shen H,Guan Q,Zhang X,等。阿尔茨海默病神经炎症的新机制:肠道菌群介导的NLRP3炎症小体的激活。Prog Neuropsychopharmacol Biol Psychiatry,2020,100:109884 [16] Ferreira-Vieira TH,Guimaraes IM,Silva FR,等。阿尔茨海默病:针对胆碱能系统。Curr Neuropharmacol,2016,14:101-15 [17] Scannevin RH。针对神经退行性蛋白质错误折叠障碍的治疗策略。Curr Opin Chem Biol,2018,44:66-74 [18] Giau VV,Lee H,Shim KH 等人。CRISPR-Cas9 的基因组编辑应用促进阿尔茨海默病的体外研究。Clin Interv Aging,2018,13:221-33 [19] Gupta D,Bhattacharjee O,Mandal D 等人。CRISPR-Cas9 系统:基因编辑的新曙光。生命科学, 2019, 232: 116636 [20] Makarova KS, Wolf YI, Alkhnbashi OS, et al.更新了