XiaoMi-AI文件搜索系统
World File Search SystemNucleus

1 基底核的神经病理学变化......
摘要 1 型或 2 型糖尿病 (T1DM 或 T2DM) 患者经常会出现认知障碍。我们对死后人脑组织中的 Meynert 基底核 (NBM) 中的细胞进行了分析,以研究神经病理学变化。71 个死后 NBM 样本按 Braak 分期 0-2 或 3-6 分为 T1DM、T2DM 和非糖尿病对照。T1DM 受试者只有 Braak 分期 0-2,因此仅与具有相似 Braak 分期的对照进行比较,而不与 Braak 分期 3-6 的受试者进行比较。我们用各自的标志物分析了表达胆碱乙酰转移酶 (ChAT)、磷酸化 Tau、神经胶质细胞和血管的神经元。我们发现与 Braak 分期 0-2 的对照和 T2DM 相比,T1DM 中 ChAT 的神经元表达明显减少。与 Braak 分期 3-6 的对照相比,T2DM 中后期过度磷酸化 Tau 水平更高。我们的结果表明,NBM 神经元乙酰胆碱生成减少可能是 1 型糖尿病患者认知功能不佳的原因。相反,2 型糖尿病可能会加剧与阿尔茨海默病样改变相关的神经病理学变化。

Newcastle PowersWapnucleus®经典
PowersWapNucleus®经典锂电系统旨在将生产力扩大到仓库/DC,生产地板和其他工业环境的所有空间中。这些坚固而轻巧的电池系统一次可每次提供8-12+小时以上的安静,无缝的AC电源。耗尽电池充电后,只需在几秒钟内用充满电的充满电的电池换成24/7操作即可。

细胞核中染色质的物理性质
基因组信息编码在长链 DNA 上,DNA 折叠成染色质并储存在微小的细胞核中。核染色质是一种带负电荷的聚合物,由 DNA、组蛋白和各种非组蛋白组成。由于其高电荷性质,染色质结构随周围环境(例如阳离子、分子拥挤等)而变化很大。过去 10 年,已经开发出捕获活细胞中染色质的新技术。我们对染色质组织的看法已从规则和静态转变为更加多变和动态。染色质形成许多紧凑的动态区域,它们充当高等真核细胞中基因组的功能单位,局部呈现液体状。通过改变 DNA 的可及性,这些区域可以控制各种功能。基于来自多功能基因组学和先进成像研究的新证据,我们讨论了拥挤的核环境中染色质的物理性质及其调控方式。

Th-229 核的激光激发
利用相干电磁辐射对基本量子系统进行共振激发是许多物理学实验的核心,例如原子和分子光谱、原子钟、量子信息处理等。相干激光激发有许多应用,特别是需要高精度控制量子叠加态的频率或相位时,但迄今为止它在核物理中几乎没有使用[1]。从典型的核激发能量和可用的激光光子能量之间的巨大不匹配可以理解激光激发原子核的困难。核激发已经在激光产生的等离子体中得到证实,其中相互作用是通过在强激光场中加速的电子介导的,电子在碰撞中或通过X射线范围内的轫致辐射与原子核相互作用[2]。不同的原子核已经通过同步辐射在6 – 60 keV能量范围内的跃迁上进行共振激发,寿命在纳秒到微秒范围内[3]。 Sc-45 的 12.4 keV 共振最近在欧洲 x 射线自由电子激光器 [4] 上被激发,其寿命为 0.47 秒。Th-229 原子核以其独特的低能同质异能态而闻名 [5 – 7] 。其激发能量为 8.4 eV,使核跃迁处于真空紫外 (VUV) 光谱范围内,使其可用于台式激光系统和精密光学工具的实验

核心寻呼站 - Repeater Builder®
本手册中的信息已经过准确性审查。但是,对于不准确的信息,我们概不负责。摩托罗拉保留对本文讨论的任何产品进行更改的权利。本文档中的信息如有更改,恕不另行通知。摩托罗拉对因本手册中的错误或遗漏而导致的硬件或软件损坏或数据丢失不承担任何责任。摩托罗拉不承担因应用或使用本文描述的任何产品或电路而产生的任何责任。摩托罗拉也不转让其专利或他人权利的任何许可。

Cochlear™Nucleus®Kanso®2声音处理器
现在可以在Nucleus Smart应用程序中获得一项新的电池健康功能,以帮助监视内置可充电电池的健康。您和您的患者将能够确定电池何时按预期运行(“好”或“好”),或者它是否需要“需要注意”或“无法充电”。当消息显示“需要注意”时,应更换电池,而不是等到消息显示“无法充电”之前。如果电池健康显示“无法充电”,您的患者可以使用便携式充电器为其声音处理器供电。如果需要,请联系您当地的人工耳蜗代表以寻求支持。
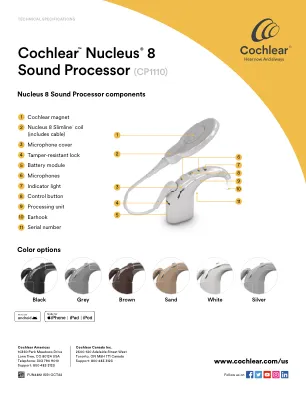
Cochlear™Nucleus®8声音处理器(CP1110)
1。人工耳蜗有限公司。D1190805。处理器尺寸比较。2022,4月*人工耳蜗建议675个锌空气电池设计用于人工耳蜗的使用。**典型的预期电池寿命是使用CI600系列或CI500系列植入物使用的默认地图设置来计算的。根据电池的年龄,每天使用的程序,植入物类型,覆盖植入物的皮肤厚度以及使用的电池大小和类型,每个用户的电池寿命各不相同。从兼容设备,真正的无线设备或FM进行流式传输可能会根据流媒体的频率和多长时间降低声音处理器电池寿命。†建议将SNR-NR,WNR和扫描提供给任何6岁及6岁以上的接收者,他们能够进行1)静态和噪音中的完整客观语音感知测试,以证明和记录性能以及2)报告对不同程序设置的偏好。††远前置只能通过听力植入专家启用。它只能为12岁及以上的用户激活,他们能够可靠地提供声音质量的反馈,并了解如何在移动不同或不断变化的环境时使用该功能。在安静的环境中使用前瞻性视频时,可能会减少语音理解。^耳蜗核8声音处理器是国际标准IEC60529的IP68的灰尘和水具有抗性。带有Aqua+的Nucleus 8 Sound处理器可以连续浸入水中,最多3米,持续2小时。始终阅读使用说明。使用Aqua+的Nucleus 8 Sound处理器是粉尘和水对国际标准IEC60529 IP68水平的抗性,当您使用耳蜗功率扩展可充电电池模块或耳蜗紧凑型电池可再充电电池模块时。参加长时间的水活动时应使用Aqua+附件。有关更多信息,请参阅相关用户指南。^^耳蜗核智能应用程序可在App Store和Google Play上找到。+耳蜗核8声音处理器与Apple和Android设备兼容。有关兼容性信息,请访问www.cochlear.com/compatibility。¥当此技术可用时,您的声音处理器将固件更新可让您连接到蓝牙LE音频兼容设备。请向您的卫生专业人员寻求有关听力损失的治疗方法的建议。结果可能会有所不同,您的健康专业人员将为您提供可能影响您结果的因素。并非所有产品都在所有国家 /地区提供。请联系您当地的人工耳蜗代表以获取产品信息。©Cochlear Limited 2022。保留所有权利。ace,偏离风格,AOS,Ardium,Autonrt,Autosositivity,Baha,Baha,Baha软件,BCDRIVE,BCDRIVE,BCDRIVE,BEAM,BEACH BATE,BATEN,BUTEN,CARINA,CARINA,CARINA,CARINA,COCHLEAR,COCHLEAR,科利耳,코클리어,코클리어,코클리어,同系,工人的软件,轮廓,轮廓,轮廓,轮廓,轮廓,自定义声音,自定义,自由,dermalock,derreferock,derrefe ofer,now,now,now,now,soy,now,soy,now,sere,now。And always, Hugfit, Human Design, Hybrid, Invisible Hearing, Kanso, LowPro, MET, MP3000, myCochlear, mySmartSound, NRT, Nucleus, Osia, Outcome Focused Fitting, Off-Stylet, Piezo Power, Profile, Slimline, SmartSound, Softip, SoundArc, True Wireless, the elliptical logo, Vistafix, Whisper, WindShield and Xidium是人工耳蜗公司的商标或注册商标。

4 月 18 日 星期四 核心大楼
Kathy Evans 博士,基因组和实验医学中心 博士生数据闪电战主席:Katy Marshall-Phelps 博士,脑科学发现中心 10.15 磷酸二酯酶 7 抑制剂:从临床前到临床开发未来成瘾症药物 Adana Keshishyan,脑科学发现中心 收入和结果:社会经济地位、早产和神经发育 Katie Mckinnon,生殖健康中心 了解 LRRK2 失调对人类干细胞衍生的星形胶质细胞的影响 Áine Heffernan,临床脑科学中心 阿尔茨海默病病理中白质的时间和区域脆弱性 Lucy Ryan,脑科学发现中心 研究自闭症儿童和青少年的睡眠问题及其与精神疾病的联系 R eesha Zahir,临床脑科学中心
核心寻呼站 - Repeater Builder®
本手册中的信息已经过准确性审查。但是,对于不准确的信息,我们概不负责。摩托罗拉保留对本文讨论的任何产品进行更改的权利。本文档中的信息如有更改,恕不另行通知。摩托罗拉对因本手册中的错误或遗漏而导致的硬件或软件损坏或数据丢失不承担任何责任。摩托罗拉不承担因应用或使用本文描述的任何产品或电路而产生的任何责任。摩托罗拉也不转让其专利或他人权利的任何许可。

癫痫脑库的概念融合了楔形核刺激的证据
许多神经系统疾病和损伤中存在的步态障碍,包括帕金森氏病(PD),中风和脊髓损伤。神经系统步态疾病在老年人中尤为常见,60岁以后的患病率超过20%(Mahlknecht等人,2013年),随着人口统计学的转变,社会健康负担可能增加。这些障碍会导致不动和跌倒,并有助于社会隔离,减少生活质量和失去独立性(Mahlknecht等,2013)。很少有治疗选择,因此在这一领域的研究中必须进行研究。在这篇观点文章中,我们回顾了导致Pedunculopontine Nucleus(PPN)进行深脑刺激(DB)的临床试验,讨论了这些试验未成功的潜在原因,并介绍了支持我们的新研究,支持我们的观点,即我们认为附近的Cune Sourture Nucleus(CNF)可能是一个更有效的目标。