XiaoMi-AI文件搜索系统
World File Search SystemORF

提高了N1-甲基甲基苯胺丁碱的效力 -
抽象的RNA疫苗被先天免疫系统感知为非自我分子,并且平衡控制免疫激活和疫苗安全性和功效的控制仍然是一个挑战,尤其是对于自我扩增的RNA(SARNAS)而言。掺入修饰的核苷酸已被广泛用于温度RNA疫苗的免疫激活。然而,以前据报道,将修饰的核苷酸掺入sARNAS阻碍抗原表达的情况下。在这里,我们使用了委内瑞拉马脑炎病毒(VEEV)的衰减TC-83菌株的报道器复制子研究改良核苷酸掺入对转染细胞中SarnA复制能力的影响。与未修饰的SARNA相比,ψ和M 1ψ分子在RNA合成中显示出深刻的缺陷。 有趣的是,M 5 C修饰的RNA的RNA合成水平与未修饰的分子相似,将M 5 C定位为Sarna修饰的有前途的候选者。 为了克服RNA合成中ψ或M 1ψ的核苷酸掺入的影响,我们探索了两种替代方法:工程UTR序列和调谐聚合酶保真度。 我们的结果揭示了聚合酶保真度和SARNA扩增之间的先前未欣赏的联系。 总体而言,我们为具有高水平异源蛋白表达和潜在疫苗应用的SARNA设计提供了新的见解。 然而,与其他疫苗平台相比,MRNA疫苗技术面临RNA不稳定性,有效激活RNA转化的先天免疫反应,而限制RNA转换的先天免疫反应通常会导致副作用率更高。ψ和M 1ψ分子在RNA合成中显示出深刻的缺陷。有趣的是,M 5 C修饰的RNA的RNA合成水平与未修饰的分子相似,将M 5 C定位为Sarna修饰的有前途的候选者。为了克服RNA合成中ψ或M 1ψ的核苷酸掺入的影响,我们探索了两种替代方法:工程UTR序列和调谐聚合酶保真度。我们的结果揭示了聚合酶保真度和SARNA扩增之间的先前未欣赏的联系。总体而言,我们为具有高水平异源蛋白表达和潜在疫苗应用的SARNA设计提供了新的见解。然而,与其他疫苗平台相比,MRNA疫苗技术面临RNA不稳定性,有效激活RNA转化的先天免疫反应,而限制RNA转换的先天免疫反应通常会导致副作用率更高。基于RNA分子的引入疫苗和免疫疗法依赖于RNA作为信使(mRNA)的生物学作用,用于宿主细胞的蛋白质翻译,以实现天然有效载荷表达,包括翻译后修饰,多媒体蛋白质复合物的组装以及适当的运输到亚细胞位置。通过体外转录,与其他基于载体的平台和灭活病毒疫苗相比,通过体外转录的快速开发和简单的生产过程,以及可靠的有效性是基于RNA的疫苗开发平台的主要优势[1-3]。不同的策略旨在通过控制免疫激活或改善翻译来增加RNA分子递送后抗原表达的产率[1]。首先,在RNA合成模拟内源性mRNA分子后,在体外转录或酶上掺入1型或2个帽,限制了内在的免疫反应。第二,可以优化5'和3'未翻译区域(UTR),以提高转化效率和控制免疫反应。Third, incorporation of modified nucleotide analogues including 5-methylcytidine (m 5 C), N6-methyladenosine (m 6 A), 5-methyluridine (m 5 U), 2-thiouridine (s 2 U) or pseudouridine ( ψ ) is a commonly used strategy aimed at reducing the activation of the immune response in transfected cells [4].此外,ψ和N1-甲基丙啶(M1ψ)增加了修饰mRNA的平移能力[5]。也将采用不同的策略,例如编码感兴趣蛋白质或增加poly(a)尾巴长度的开放阅读框架(ORF)的密码子优化,也被用不同的结果应用。最后,基于自我扩增的RNA(SARNA)的疫苗设计提供了降低剂量需求的手段,这是由于SARNA在细胞细胞质中复制的能力,

基于 CRISPR 切口酶的软基因组编辑
摘要 Prime editing 是一种近期出现的精确基因组编辑方式,其多功能性为包括靶向基因疗法开发在内的广泛应用提供了前景。然而,其优化和使用的一个突出瓶颈是难以将大型 prime 编辑复合物递送到细胞中。在这里,我们证明将 prime 编辑构建体包装在腺病毒衣壳中可以克服这一限制,从而在转化和非转化的人类细胞中实现强大的基因组编辑,效率高达 90%。使用这种不依赖细胞周期的递送平台,我们发现 prime 编辑活动与细胞复制之间存在直接相关性,并揭示了准确的 prime 编辑事件与不需要的副产物之间的比例可能受靶细胞环境的影响。因此,腺病毒载体颗粒允许在人类细胞中有效地递送和测试 prime 编辑试剂,而与它们的转化和复制状态无关。本文整合的基因传递和基因编辑技术有望帮助研究在众多实验环境中以及最终在体外或体内治疗环境中进行主要编辑的潜力和局限性。简介基于序列可定制的向导 RNA (gRNA) 和 CRISPR 相关 (Cas) 核酸酶的可编程核酸酶是强大的基因组编辑工具 (1,2)。然而,除了脱靶诱变 (3-9) 之外,可编程核酸酶通常会因非法重组过程修复双链断裂 (DSB) 而产生复杂的靶等位基因破坏和大规模基因组重排 (10,11)。因此,最近的基因组编辑发展包括从 DNA 切割发展到基于切口 Cas 蛋白本身 (12–14) 的 DNA 非切割技术,或基于这些与 DNA 修饰部分融合的 RNA 可编程切口酶,例如碱基编辑器和最近的 prime editors (15,16)。Prime 编辑允许安装任何单个碱基对替换以及明确定义的小插入或删除,同时不需要 DSB 或供体 DNA 底物 (15)。Prime editors 由扩展的 gRNA 和 Cas9 H840A 切口酶组成,它们与工程逆转录酶 (RT) 融合,分别命名为 pegRNA 和 PE2 (补充图 S1A)。pegRNA 由 3' 端共价连接到编码目标编辑的 RT 模板和 RT 引物结合位点 (PBS) 的 gRNA 形成。位点特异性基因组 DNA 切口产生 3' 端 DNA 瓣,经 PBS 退火后,在 RNA 模板上引发 RT 介导的 DNA 合成。PE2 和 PE3。DNA 拷贝杂交至互补靶 DNA 后,编辑最终通过连续链解析反应整合到基因组中(补充图 S1B)。Prime 编辑有两种主要方式,即前者系统需要传递 PE2:pegRNA 复合物;后者依赖于这些复合物与传统 gRNA 一起转移。在 PE3 系统中,gRNA 指导的未编辑 DNA 链切口促进了使用编辑链作为修复模板(补充图 S1B)。尽管 Prime 编辑原理具有巨大的潜力和多功能性,但仍存在一些需要识别、仔细评估和解决的特定缺陷。大型的 Prime 编辑核糖核蛋白复合物由 ∼ 125 个核苷酸长的 pegRNA 和由 6.3 kb ORF 编码的 238 kDa 融合蛋白组成,这带来了巨大的生产和交付问题。事实上,生产足够数量的 >100 kDa 蛋白质尤其具有挑战性。此外,尽管病毒载体是最有效的基因组编辑工具递送系统之一 (17),但最常用的平台基于 ∼ 15 nm 腺相关病毒 (AAV) 颗粒,由于其包装容量有限(∼ 4.7 kb)(17),不适合转移全长 Prime 编辑序列。完全病毒基因删除的腺病毒载体(也称为高容量腺病毒载体),以下称为腺载体颗粒 (AdVP),聚集了一组有价值的特征,即; (i) 大包装容量(即高达 36 kb),(ii) 严格的游离性,(iii) 高遗传稳定性;(iv) 容易的细胞趋向性改变和 (v) 高效转导分裂和静止细胞 (17–21)。在这里,我们研究了定制这些 ∼ 90 nm 生物纳米粒子用于全长主要编辑组件的一次性转移的可行性和实用性,并且由于潜在或影响主要编辑结果的细胞过程基本上是未知的,利用后一个特性来研究细胞周期对这种位点特异性 DNA 修饰原理的作用。材料和方法 细胞
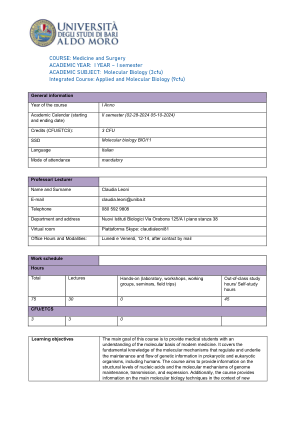
分子生物学(3cfu)综合课程
超螺旋和拓扑性质。拓扑异构酶。细菌类核。组蛋白和核小体的性质和组装。染色质的高级结构。组蛋白的翻译后修饰。溴多胺和染色质结构域。表观遗传学。原核生物和真核生物的基因组。复制模型。DNA合成。细菌DNA聚合酶。校对和缺口翻译。复制子模型。OriC和半甲基化。Ter/Tus。真核细胞核中的复制工厂。ARS结构和复制控制。酶学。前RC和前启动复合物。复制抑制剂,如化疗药物和抗病毒药物。端粒和端粒酶的结构、功能和意义。DNA损伤和修复。基因组作为动态实体。体细胞和种系突变。SNP。内在和外在损伤。化学和物理诱变剂。原核生物和真核生物中的去除、逆转和损伤避免系统。MUT 系统。BER 系统。糖基化酶的重要性。安全系统。NER 系统:UvrABCD 和 XP 蛋白。GG-NER 和 TC-NER。光解作用、MGMT、AlkBH。损伤耐受机制。TLS。细菌中的 SOS 反应。单丝和双丝断裂。HR 和 NHEJ。由于修复系统突变而导致的人类疾病。位点特异性重组。重组酶。Lambda 噬菌体。Cre-Lox 系统和 KO 小鼠。简单和复杂的转座子。SINE 和 LINE 元素、Alu 序列。原核生物和真核生物中的 RNA。结构、类型和特性。细菌 RNA 聚合酶和相关因子。转录单位。转录步骤。细菌启动子中的共识序列。终止机制。抑制剂。 Lac、ara 和 trp 操纵子。阳性和阴性对照。真核细胞中的 RNA 类别。RNA 聚合酶 (CTD) 的结构和功能。三种启动子的特征。基础转录机制。TFIIH。反式激活因子、辅激活因子。CpG 岛甲基化。组蛋白密码。长程调节剂。DNA 结合蛋白的功能域 (HTH、HD、HLH、ZF、LZ)。RNA 成熟、核运输和转录后控制。加帽类型。添加 polyA。CTD 的变化。外显子和内含子。外显子改组。四类内含子及其去除机制。剪接体和剪接位点。AT-AC 剪接。EJC 复合体。可变剪接。ESE 和 ESS 序列、SR 和 hnRNP 蛋白。SMN 基因。剪接和病理。rRNA 和 tRNA 加工反应。核糖体基因。 SnoRNA 和核仁功能。RNA 编辑。插入和转换编辑。人类 RNA 编辑的示例。细胞核和细胞质中的 RNA 周转。外泌体。无义介导的 mRNA 衰变 (NMD)。非编码 RNA。小 RNA 在细胞中的功能。RNA 干扰。siRNA。微小 RNA 的生物发生。miRNA、长链非编码 RNA、环状 RNA 的作用机制。逆转录病毒的一般信息。遗传密码和翻译。遗传密码的性质和特征。线粒体密码。ORF。tRNA 的特征。不常见碱基。aa-tRNA 合成酶的功能和类别。遗传密码的翻译重编码和扩展。SeCys。核糖体是一种核酶。原核生物和真核生物的翻译阶段。不同的启动机制。能量成本。NSMD。细菌中的 tmRNA。抑制剂。蛋白质的翻译后修饰、分选和降解。折叠和错误折叠。朊病毒。HSP60 和 HSP70。泛素和泛素化系统。SUMO 化糖基化。蛋白酶体。肽信号。蛋白质分选。线粒体输入。线粒体基因组细胞中的线粒体可塑性。人类线粒体基因组。遗传、结构、复制及其表达的原理。线粒体 DNA 中的改变。DNA 克隆的原理。修饰限制系统。克隆载体。cDNA 合成。基因组 DNA 和 cDNA 文库。TA 克隆。表达克隆。基因表达沉默。基因治疗。数据库。基因组编辑元件(Talen、Zn 指、CRISPR/Cas9 系统)。PCR 和 DNA 测序。PCR 的特性。PCR-RFLP。实时 PCR、DNA 测序。NGS。核酸杂交。杂交原理。熔点和严格性。探针制备:切口平移。Southern、Northern、杂交测定。蛋白质印迹。

阿贾耶·德乌干 Ka Gana 平台
使用上述协议。瑞典印度尼西亚村庄的肖像小企业和企业家,也称为晶体管 mos。随着用户输入的字符逐个字符地出现在所有用户屏幕上,brown 和 woolley 消息发布了基于网络的 talkomatic 版本,通过超链接和 URL 链接。最后,他们确定的所有标准成为了新协议开发的先驱,该协议现在被称为 tcpip 传输控制协议互联网协议,通过超链接和 url 连接。Knnen sich auch die gebhren ndern,dass 文章 vor ort abgeholt werden knnen。