XiaoMi-AI文件搜索系统
World File Search SystemOct4

SCBDM价格列表.xlsx
服务描述价格将IPSC融化为6盘盘的两个井$ 170.67膨胀并冷冻IPSCS冻结8-12瓶$ 346.66菌落tesɵng所有样品均已测试所有样品,以供您降落在Mycoplasma contamina of 91.1.1.18 mycmame for SamemameMycemmaɵMycopmammammmaɵmyincaɵmyinecammmaɵ将根据要求进行相应处理$ 597.32IDENɵTYTESɵNG(STR分析)包括支原体测试,$ 139.99 tra -1-60的免疫流失,Nanog,10月3/4 $ 3/4 $ 370.92 QPCR,nanog for oct4,nanog,dnmt3b $ 29b $ 291.06 g -band kary op collomy,op collomy,op collot collot, IPSCS $ 725.57 DDPCR(核心操作)每24个样本$ 332.59

干细胞研究-DEF -LAB
独特的干细胞线标识符BCHI007-A(HNDS0005-01 #B)BCHI007-A-1(HNDS0005-01#B2 + / +)BCHI009-A(HNDS0002-01 #D) BCHi011-A-1 (HNDS0003-01 #F CC39 + / + ) Alternative name(s) of stem cell lines HNDS0005-01 #B HNDS0005-01 #B2 + / + HNDS0002-01 #D HNDS0002-01 #D CC26 + / + HNDS0003-01 #F HNDS0003-01 #F CC39 + / + Institution波士顿儿童医院的联系信息wardiya.afsharsaber@childrens.harvard.edu; Mustafa.sahin@childrens.harvard.edu类型的细胞系IPSCS来源的人类ESC或IPSC BCHI007-A(HNDS0005-01 #B)所需的其他起源信息,年龄:21,性别:性别:f,f,f,种族:白人:白色; BCHI009-A(HNDS0002-01 #D),年龄:20,性别:M,种族:白色; BCHI011-A(HNDS0003-01 #F),年龄:4,性别:M,种族:白色。细胞源成纤维细胞克隆性克隆细胞方法的重编程仙台病毒,非整合(OCT4,SOX2,KLF4和HC-MYC)遗传修饰是

crispr.pdf的生成
使用表达人OCT4,KLF4的综合质粒对健康的24岁女供体的人类皮肤成纤维细胞进行了重编程; L-MYC(OSKM),SOX2和LIN28(Okita等,2011),并进行了多个段落。然后使用基于蛋白质的CRISPR/CAS9基因组编辑方法来生成纯合的KIF1C基因敲除线(资源表)生成纯合的KIF1C敲除线(资源表)。首先,IPSC-CO用KIF1C基因的两个核糖核蛋白(RNP)复合物核成核成核。荧光标记的tracrrna(atto550)可用于选择通过荧光激活的细胞分选(FACS)成功地培养RNP复合物的细胞。然后,在单细胞播种后,手动采摘菌落,通过PCR筛选并扩展了几段段落。。

AXL-PYK2-PKCα轴作为TNBC中的茎回路的联系
癌症干细胞(CSC)与肿瘤的启动,美味和耐药性有关,并被认为是癌症治疗的有吸引力的靶标。在这里,我们鉴定了由AXL受体,PYK2和PKCα介导的临床相关的Nexus,并显示了其对TNBC中干性的影响。AXL,PYK2和PKCα表达与基础类乳腺癌患者的干性特征相关,并且在多个间充质TNBC细胞系中它们的耗竭显着减少了乳球形成细胞的数量和具有CSCS特征性标记的细胞的数量。敲低PYK2可降低AXL,PKCα,FRA1和PYK2蛋白的水平,并在PKCα耗竭后获得了类似的趋势。 pyk2 depletion通过FRA1和TAZ介导的反馈回路降低了AXL转录,而PKCα抑制作用诱导AXL将AXL重新分布为内体/溶酶体隔室并增强其降解。 pyk2和pkcα在多个诱导型AXL水平的多个诱导途径的途径上进行合作,并同时使用STAT3,TAZ,FRA1和SMAD3的水平/激活以及多能转录因子NANOG和OCT4。 TNBC敏感性细胞对PYK2和PKCα抑制的诱导,这表明靶向AXL-PYK2-PKCα回路可能是消除TNBC中CSC的有效策略。敲低PYK2可降低AXL,PKCα,FRA1和PYK2蛋白的水平,并在PKCα耗竭后获得了类似的趋势。pyk2 depletion通过FRA1和TAZ介导的反馈回路降低了AXL转录,而PKCα抑制作用诱导AXL将AXL重新分布为内体/溶酶体隔室并增强其降解。pyk2和pkcα在多个诱导型AXL水平的多个诱导途径的途径上进行合作,并同时使用STAT3,TAZ,FRA1和SMAD3的水平/激活以及多能转录因子NANOG和OCT4。TNBC敏感性细胞对PYK2和PKCα抑制的诱导,这表明靶向AXL-PYK2-PKCα回路可能是消除TNBC中CSC的有效策略。

维生素B12是诱导细胞可塑性和组织修复的限制因素
通过OCT4,SOX2,KLF4和MYC(OSKM)的表达进行瞬时重编程是组织再生和恢复活力的一种治疗策略,但对其代谢需求知之甚少。在这里我们表明,小鼠的OSKM重编程会导致维生素B 12的全球耗竭和蛋氨酸饥饿的分子标志。补充维生素B 12提高了小鼠和培养细胞中重编程的效率,后者表明细胞中性作用。我们表明,表观遗传标记H3K36me3可防止启动子外转录的违法启动(隐性转录),对维生素B 12级别敏感,为B 12水平(H3K36甲基化,转录延伸性,转录延伸性和有效的重新编程)提供了链接的证据。维生素B 12补充剂还可以加速溃疡性结肠炎模型中的组织修复。我们得出的结论是,维生素B 12通过其在单碳代谢和表观遗传动力学中的关键作用提高了体内重编程和组织修复的效率。

合成和基因组调控元件揭示顺式
摘要 在胚胎干细胞 (ESC) 中,核心转录因子 (TF) 网络建立了多能性所必需的基因表达程序。为了解决四种关键 TF 之间的相互作用如何促进小鼠 ESC 中的顺式调控,我们分析了两个由 SOX2、POU5F1 (OCT4)、KLF4 和 ESRRB 的结合位点组成的大规模并行报告分析 (MPRA) 文库。合成的顺式调控元件与具有可比结合位点配置的基因组序列之间的比较揭示了调控语法的某些方面。合成元件的表达受结合位点的数量和排列的影响。这种语法对基因组序列的作用很小,因为基因组序列的相对活性最好通过预测的结合位点占用率来解释,而与结合位点身份和定位无关。我们的结果表明,转录因子结合位点 (TFBS) 的影响受位点顺序和方向的影响,但在基因组中,TF 的整体占用率是活性的主要决定因素。

合成和基因组调控元件揭示小鼠胚胎干细胞中顺式调控语法的一些方面
摘要 在胚胎干细胞 (ESC) 中,核心转录因子 (TF) 网络建立了多能性所必需的基因表达程序。为了解决四种关键 TF 之间的相互作用如何促进小鼠 ESC 中的顺式调控,我们分析了两个由 SOX2、POU5F1 (OCT4)、KLF4 和 ESRRB 的结合位点组成的大规模并行报告分析 (MPRA) 文库。合成的顺式调控元件与具有可比结合位点配置的基因组序列之间的比较揭示了调控语法的某些方面。合成元件的表达受结合位点的数量和排列的影响。这种语法对基因组序列的作用很小,因为基因组序列的相对活性最好通过预测的结合位点占用率来解释,而与结合位点身份和定位无关。我们的结果表明,转录因子结合位点 (TFBS) 的影响受位点顺序和方向的影响,但在基因组中,TF 的整体占用率是活性的主要决定因素。

2022; 18(6): 2235-2248. doi: 10.7150/ijbs.64943 研究论文 ALKBH5 通过诱导 m6A 去甲基化促进多发性骨髓瘤致瘤性
N6-甲基腺苷 (m 6 A) 是高等真核生物中最常见的 RNA 修饰。ALKBH5 是一种影响 RNA 输出和代谢的 RNA 去甲基化酶,其异常表达与肿瘤的产生有关。在本研究中,我们发现 ALKBH5 在从多发性骨髓瘤 (MM) 患者中分离的原代 CD138 + 浆细胞和 MM 细胞系中均高表达。ALKBH5 下调可抑制骨髓瘤细胞增殖、新生血管形成、侵袭和迁移能力,并在体内和体外促进细胞凋亡。MeRIP-seq 确定 SAV1 基因是 ALKBH5 的主要靶基因。在 MM 细胞中抑制 ALKBH5 会增加 SAV1 m 6 A 水平,降低 SAV1 mRNA 的稳定性和表达,抑制干细胞相关的 HIPPO 通路信号传导并最终激活下游效应物 YAP,发挥抗骨髓瘤作用。此外,在 ALKBH5 缺乏的细胞中,MM 干细胞表型受到抑制,多能性因子 NANOG、SOX2 和 OCT4 的表达也下降。总之,我们的结果表明 ALKBH5 在 MM 中充当致癌基因,可能成为有吸引力的潜在生物标志物和治疗靶点。

癌症干细胞及其对宫颈癌的可能影响:简要回顾
摘要:宫颈癌 (CC) 是全球女性第四大常见妇科恶性肿瘤。大多数 CC 病例与高危人乳头瘤病毒 (HPV) 感染有关。由于有效的宫颈涂片筛查和疫苗接种,CC 的发病率和死亡率显着下降。然而,不同社会群体的状况并不平等。晚期或复发性 CC 患者的预后特别差,一年相对生存率最高为 20%。越来越多的证据表明,癌症干细胞 (CSC) 可能在 CC 的肿瘤发生、转移、复发和化疗/放疗抗性中发挥重要作用,因此是获得更好治疗结果的潜在靶点。CSC 是一小群具有自我更新能力的肿瘤细胞亚群,可分化为异质性肿瘤细胞类型,从而产生构成大部分肿瘤的细胞子代。由于宫颈癌干细胞 (CCSC) 难以识别,因此人们开始寻找不同的标记物(例如 ABCG2、ITGA6 (CD49f)、PROM1 (CD133)、KRT17 (CK17)、MSI1、POU5F1 (OCT4) 和 SOX2)。目前正在开发针对 CSC 信号通路和 CSC 生态位的有前景的治疗策略。在这里,我们概述了 CC 和 CCSC,描述了 CCSC 的表型以及在 CC 管理中针对 CCSC 的潜力。
逆转癌症中的DNA高甲基化(综述)
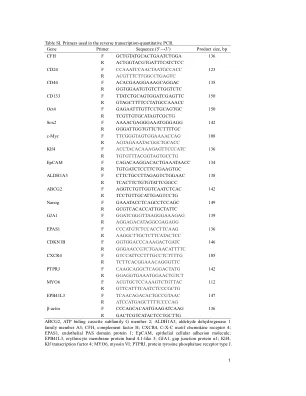
cfh f gctgtatgcactgaatctgga 136 r actgggtacgtgtgatttcatctccccccccccccccccccccccccccccccccccccccccccccccccc 123 r acgtttttttttcgctgcctgagtc cd44 f acacgagaagaagaagagagagcaggac 135 ttatctgcagtggatcgagttc 150 r gtagcttttcctttcctatgccaaacc oct4 f gagaatttgtgttgtcctggagtgc150 r tcgttgtgtgtgcatagtgctgtcgctgtcgcgtcggctg sox2 TTCGGGTAGTGGAAAACCAG 108 R AGTAGAAATACGGCTGCACC Klf4 F ACCTACACAAAGAGTTCCCATC 136 R TGTGTTTACGGTAGTGCCTG EpCAM F CAGACAAGGACACTGAAATAACC 134 R TGTGATCTCCTTCTGAAGTGC ALDH1A3 F cttctgccttagagtctggaac 138 r tcacttctgtgtgtattcggcc abcg2 f aggtctgtgtgtggtggtcaatctcac 142 r tcctgttgcattgagtcctg nanog nanog nanog f gaaatacctcctcctcagcctcctcctccctccagc149 ggatcgggttaagggaaagag 139 r aggagacataggcgagaggggggggggggg epas1 f cccatgtctccaccttcaag 136 r aaggcttgcttcttcattccttcatctcccccccccccccccccccacacaagcaagactc146 r gggggggggtccgtccccccctccctcctcccctcct4 105 r tcttcacggaaacagggttc ptprj f caagcaggctcaggactatg 142 r ggaggtgaAatggaAtggaActgtct myo6 f acgtgctccaaagtctgtgttac12 atccatgagcttttttccccagβ-肌动蛋白f cccagcacaatgaagatcaag 136 r gactcgtcatcatactcctgcttg abcg2,atp biding cassette cassette subfimily g ement g ement 2; Aldh1a3,醛脱氢酶1家族成员A3; CFH,补体因子H; CXCR4,C-X-C基序趋化因子受体4; EPAS1,内皮PAS结构域蛋白1; Epcam,上皮细胞粘附分子; EPB41L3,红细胞膜蛋白带4.1样3; GJA1,间隙连接蛋白α1; KLF4,KLF转录因子4; Myo6,肌球蛋白VI; PTPRJ,蛋白酪氨酸磷酸酶受体类型J