XiaoMi-AI文件搜索系统
World File Search SystemOlsen

EIB世界贸易头条
匈牙利国民因密谋向俄罗斯政府最终用户出口美国军事级收音机而被捕 - 2024年8月26日,司法部未公开刑事诉讼,要求刑事申诉,指控匈牙利民族本德·霍尔瓦斯(Hungarian National Bense Horvath)违反了针对俄罗斯的美国出口控制,包括与其他人合谋非法出口美国 - 美国 - 原始批准的无线电通讯技术,而无需向俄罗斯俄罗斯的最终用户出口。horvath被投诉指控一项密谋违反《 2018年出口控制改革法案》(ECRA)。他于8月23日到达加利福尼亚州旧金山的旧金山国际机场时被捕。“正如所谓的,被告试图使用跨国采购链为俄罗斯实体购买军事级收音机,以逃避执法部门,”司法部国家安全部门的助理总检察长马修·奥尔森(Matthew G. Olsen)说。“司法部仍致力于破坏和持有负责任的犯罪网络,这些犯罪网络继续推动国外的俄罗斯侵略并威胁我们的集体安全。”商务部工业与安全局(BIS)的出口执法助理秘书Matthew S. Axelrod表示:“针对为俄罗斯政府装备俄罗斯政府的非法全球采购网络是为俄罗斯政府配备俄罗斯政府的最高优先事项。”投诉称,Horvath本人安排通过位于西班牙,塞尔维亚,匈牙利,拉脱维亚和其他地方的一个分支机构网络购买美国 - 皇家无线电通信技术,并将这种技术走私到俄罗斯政府最终用户。“正如Horvath的逮捕所表明的那样,在您经营的世界上,当美国认为您的行为违反我们的出口法时,我们采取行动都没关系。”美国检察官Matthew M. Graves在哥伦比亚特区说:“据称,这名被告试图绕过美国的出口管制,以保护我们的国家安全,并解决俄罗斯无端的全面入侵乌克兰。”“我们将继续与我们的合作伙伴合作,将计划确保美国技术违反美国法律的人绳之以法。”根据法院文件,Horvath和其他人管理了一个跨国采购网络,该网络直接与俄罗斯政府的各个实体签约,并从事大规模项目,例如在俄罗斯/乌克兰边境俄罗斯库尔斯克地区建造运营无线电通信系统。至少从2023年1月左右开始,Horvath及其网络中的其他人与一家美国小型广播分销公司进行了讨论,涉及采购和出口到俄罗斯美国制造的军事级收音机和相关配件。在接下来的几个月中,Horvath继续努力确保这些物品,他打算通过拉脱维亚的货运代理转移到俄罗斯。

长寿生物技术:连接人工智能、生物标志物、老年科学和临床应用,实现健康长寿
Yu-xuan Lyu 1,2,* , qiang fu 3,4,* , dominic wick 6,125,* , kejun ying 7,* , Aaron King Kaya 13 , Andrea B. Maier 14 , Andrea Olsen 15 , Anja Groth 16 , Anna Katharina Simon 17,18 , Anne Brunet 19 , Aisyah Jamil 20 , Anton Kulaga 22 , Benjamin Yaden Örnumacher 25 , Boris DjordJervic 26,27 , Brian Kennedy 14 , Chieh Chen 28,29 , Christine Yuan Huang 30 , Christopph U. Correll 31,32 , Collin y. , Dariusz Sołdacki 40 , David Erritzoe 41 , David Meyer 25 , Sinclair 42 , Eduardo Nunesni 43 , Emma C. Teeling 48 , Evandro F. Fang 49 , Evelyne Bischof 50 , Evi M. Mercken 51 , Fabian Finger 52 , Folkert Kuipers , Frank W. Pun 54 , Gabor Gyünze , Gari Harold A. Pincus 59 , Joshua McClure 60 , James L. Kirkland 61 , James Peyer 62 , Jamie N. Justice 63 , Jan VIJG 64 , Jennifer R. Gruhn 65 , Jerry mlaughlin 66 , Joan Mannick , Joe Betts-Lacroix 70 , John M. Sedivy 71 , John R. Speakman 72 , Jordan Shlain 73 , Julia von Maltzahn 74 , Katrin I. Andreasson 75 , Krikaras fort 76 , Constantnus Palikaras for Feer 78 , Lene Juel Rasmussen 79 , Liesbeth M. Veenhoff 53 , Lisa Melton 80 , Luigi ferrucci 81 , Marco Quarta 82,83,84 , Maria Kval 85 , Maria Marinova 86 , Mark Gingel 89 , Milos Filipovic 90 , Mourad Topors 91 , Nataly Mitin 92 , Nawal Roy 93 , Nika Pintar 94 , NIR BARZILAI , ter O. Fedichev 98 , Petrina Kamya 99 , Pura Muñoz-Canoves 100 , Rafael de Cabo 101 , Richard Garagher 102 , Rob Konrad 103 , Roberto ripa 2 , Sabrina Bütttttttttttttttttttttttttttnner , Sebastian Brumeeier 107 , Sergey Jakimov 57 , Shan Luo 108 , Sharon Rosenzweig-Plipson 66 , Shih-Yin Tsai 109 , Stefanie Dimmeler 110 , Thomas R. , Tony Wyss-Coray 75 , toy finel 115 , tzispora strauss 116,117 , Vadyshev 7 , Valter D. song. Zo Sorsinino 14 , Vittorio Sebastiano 122 , Wenbin Li 123 , Yousin Suh 124 , Alex Zhavoronkov 20 , Morten Scheeketee-Knudensen 79 , Daniela Bakula

攀岩专项运动测试:能量系统贡献及其与运动表现的关系
由于运动攀岩越来越受欢迎,它被列入 2020 年奥运会比赛项目。此外,过去 30 年,在岩石上而非比赛期间取得的最高攀岩成绩呈渐近式增长 ( Michailov, 2014 )。这两个事实都表明攀岩运动已经达到了发展的高级阶段。这对攀岩者的准备工作提出了更高的要求,需要监测和评估攀岩特定的体能,以优化训练并进一步提高攀岩表现。事实证明,传统使用的锻炼测试对于评估攀岩者的训练状态毫无用处 ( Watts, 2004 )。为了为攀岩者选择合适的锻炼测试,应该熟悉攀岩中的特定负荷特性、表现限制因素和生理方面。攀岩运动种类繁多,持续时间和锻炼强度各不相同。比赛期间,先锋攀登的时间限制为 6 分钟。否则,运动攀登路线(领先)的上升通常需要 1-4 分钟(红点 - 制定路线后)和 3-10 分钟(即兴攀登 - 首次尝试)。抱石攀登通常持续 30-50 秒(Michailov,2014)。在抱石比赛期间,攀岩者可以根据需要多次尝试抱石问题,并且可以在 4 到 5 分钟内完成。之后,他们休息 4 到 5 分钟,然后开始处理下一个抱石问题。男子 15 米速度攀登的实际记录是 5.21 秒。因此,攀登并不等同于永久性最大努力,而是多种肌肉努力模式的混合,由与最大力量相关的收缩强度、收缩阶段的持续时间及其与放松阶段的关系决定。所有攀岩运动的共同点是,它们需要剧烈的间歇性等长肌肉收缩(Sheel,2004)。手指屈肌的收缩时间比其放松时间长得多。收缩与放松的比率会限制血流。在运动攀岩中,该比率可能为 4:1,在抱石运动中,该比率可能为 13:1(Schadle-Schardt,1998;White and Olsen,2010)。攀岩表现的结构包括大量运动能力和技能,包括生理和心理因素、人体测量特征和灵活性(Sheel,2004;Watts,2004;Giles 等,2006;Michailov,2014)。身体变量在很大程度上解释了攀岩表现的差异,这些变量是可训练的因素,例如手指手臂的力量和耐力,而人体测量特征和灵活性的影响相对较小(Mermier 等人,2000 年;Baláš 等人,2012 年;Laffiaye 等人,2016 年)。身体、技术和心理特征以类似的方式解释了攀岩表现的结构,这可以作为攀岩者需要进行和谐发展训练的证据(Magiera 等人,2013 年)。从生理学的角度来看,攀岩是一项有趣的运动,因为它需要:(a)令人满意的有氧能力和一般耐力,以及(b)由有氧、磷酸原[三磷酸腺苷(ATP)和磷酸肌酸(PCr)]和无氧乳酸能量系统提供的特定肌肉力量和耐力(Sheel,2004;Watts,2004;Giles 等人,2006;Bertuzzi 等人,2007)。先前的研究重点是

利用CRISPR/Cas9进行基因组编辑的原理与过敏性疾病......
生物化学研究 2008 : 63 : 17 ― 20. 5) Carroll D. 利用可靶向核酸酶进行基因组工程。生物化学年鉴2014; 83:409―39.6)Jinek M、Chylinski K、Fonfara I、Hauer M、Doudna JA、Charpentier E. 适应性细菌免疫中的可编程双RNA引导DNA内切酶。科学 2012; 337:816―21.7)Gasiunas G、Barrangou R、Horvath P、Siksnys V. Cas9-crRNA 核糖核蛋白复合物介导特异性 DNA 切割以实现细菌适应性免疫。美国国家科学院院刊2012; 109:E2579―86. 8) Nakata A,Shinagawa H,Amemura M.大肠杆菌碱性磷酸酶同工酶基因(iap)的克隆。基因 1982; 19: 313 -- 9. 9) Nakata A、Amemura M、Makino K. 大肠杆菌 K-12 染色体中重复序列的异常核苷酸排列。细菌学杂志1989; 171: 3553 ― 6.10) Groenen PM、Bunschoten AE、van Soolingen D、van Embden JD。结核分枝杆菌直接重复簇中 DNA 多态性的性质;通过一种新颖的分型方法进行菌株鉴别的应用。分子微生物学1993; 10: 1057 — 65。11) Mojica FJ、Judge G、Rodriguez-Valera F. 不同盐度下邻近部分修饰的 PstI 位点的 Haloferax medi- terranei 序列的转录。分子微生物学1993; 9:613―21。12)Bult CJ,White O,Olsen GJ,Zhou L,Fleischmann RD,Sutton GG 等。产甲烷古菌 Methanococcus jannaschii 的完整基因组序列。科学 1996 ; 273: 1058 ― 73.13) Haft DH,Selengut J,Mongodin EF,Nelson KE。原核生物基因组中存在 45 个 CRISPR 相关 (Cas) 蛋白家族和多种 CRISPR/Cas 亚型。 PLoS Comput Biol 2005; 1:e6 14) Makarova KS、Aravind L、Grishin NV、Rogozin IB、Koonin EV。通过基因组背景分析预测的嗜热古菌和细菌特有的 DNA 修复系统。核酸研究2002; 30:482―96.15)Makarova KS,Aravind L,Wolf YI,Koonin EV。 Cas 蛋白家族的统一以及 CRISPR-Cas 系统起源和进化的简单场景。直接生物学2011; 6:38。16) Mojica FJM、Ten-Villaseñor C、Garcia-Martinez J、Soria E. 间隔规则的原核重复序列的介入序列源自外来遗传元素。 J Mol Evol.2005; 60: 174 ― 82。17) Pourcel C、Salvignol G、Vergnaud G. 鼠疫耶尔森氏菌中的 CRISPR 元素通过优先吸收噬菌体 DNA 获得新的重复序列。微生物学 2005; 151: 653 ― 63.18) Bolotin A, Quinquis B, Sorokin A, Ehrlich SD。

应对传染病重大挑战的新机遇
历史上,传染病给人类带来了沉重的打击。历史一再警告我们,一种致命的病原体就能杀死数百万人。14 世纪席卷欧亚大陆的黑死病大流行夺走了多达 1 亿人的生命( Cohn,2008 ),1918 年的西班牙流感在不到 2 年的时间内夺走了 5000 多万人的生命( Taubenberger and Morens,2019 )。这种情况在 20 世纪开始发生变化,抗生素和疫苗这两项了不起的成就拯救了数亿人的生命,使他们免于致命感染。如果我们没有针对天花、黄热病、脊髓灰质炎和其他致命病原体的疫苗,难以想象会有多少人丧生。如果我们没有抗生素,外科病房会发生什么情况则令人难以想象。一个令人愉快的巧合是,导致这些巨大成功的工具和技术往往是由微生物本身提供的:抗生素是由细菌和真菌产生的,疫苗通常是减毒或灭活的微生物。同样令人着迷的是,包括病毒和细菌在内的微生物教会了我们分子语言,让我们理解生命最基本的过程,并启发我们开发强大的生物技术来预防和治疗各种危及生命的感染。现代健康科学的一个支柱是 DNA 生物学和重组 DNA 技术。正是细菌和病毒教会了我们 DNA 是遗传物质,以及 DNA 基因表达是如何执行和调控的。更值得庆幸的是,我们还从这些微生物那里获得了解码 DNA 序列和设计 DNA 克隆的分子工具。如今,下一代测序和元数据分析彻底改变了我们在诊断、预防和治疗层面管理传染病的方式。尽管取得了这些突破性的成就,但传染病仍然给公共卫生带来沉重的负担,每年造成 1000 万至 1500 万人死亡。为证明这一严重的全球影响,世界卫生组织 (WHO) 于 2019 年公布的全球十大健康威胁中有六项与传染病有关 (https://www.who.int/emergencies/ten-threats-to-global-health-in-2019)。这六大威胁包括全球流感大流行、抗生素耐药性、埃博拉和其他高威胁病原体、疫苗犹豫、登革热和艾滋病毒 (HIV)。这些传染性病原体和相关问题位列全球卫生挑战之首并非偶然。人类历史上经常发生流感疫情。我们根本无法从人类中根除流感病毒,部分原因是它们会从鸟类和其他动物的天然宿主偶尔传播给人类 (Olsen 等人,2006 年)。生产有效的季节性流感疫苗已经是一个挑战,这将是一项更加艰巨的任务,预测和准备应对不可预测但即将来临的流感大流行,这在目前并非不可能。几十年来,我们一直受益于抗生素的使用。然而,过度使用抗生素和其他不良医疗习惯加速了耐药细菌的出现。如果没有可持续的新抗生素渠道,也没有其他有效的细菌感染治疗方法,我们可能会死于多重耐药致病菌(也称为超级细菌)引起的感染。据美国疾病控制和预防中心报道,仅在美国,每年就有 35,000 人死于抗生素耐药性细菌感染。
HBB 追踪.xlsx
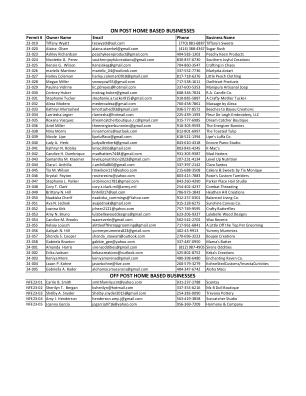
许可证编号 所有者姓名 电子邮件 电话 公司名称 23-019 Tiffany Wyatt tecwyatt@aol.com (770) 881-6887 Tiffany's Sweets 23-020 Alaina Olsen alaina.staerkel@gmail.com (414) 388-4367 Sugar Rush 23-023 Ashley Richardson peachykeenproduct@gmail.com 404-583-1303 Peachy Keen Products 23-024 Nicolette G. Perez southernjoyfulcreations@gmail.com 830-837-6730 Southern Joyful Creations 23-025 Kenzie G. Wilson Kenziekeg@gmail.com 704-860-3547 Crafting in Chaos 23-026 marieliz Martinez marieliz_04@outlook.com 337-552-7736 Marlydia.dotart 23-027 Harley Coleman harley.coleman0918@gmail.com 817-718-6376 Little Peach Clothing 23-028 Megan Miller snowyowl55@gmail.com 717-538-1611 OwlHeart Products 23-029 Paulina Vidrine lic.p0reyes@hotmail.com 337-600-5353 Manipura Artisanal Soap 23-030 Cortney Huber mckrag.huber@gmail.com 608-346-7824 RA Candle Co. 23-031 Stephanie Tucker stephanie.a.tucker915@gmail.com 910-885-5887 A Crafty Mother Tucker 23-032 Alexa Modero mederoalexa@gmail.com 760-458-7861 Massage by Alexa 23-033 Kathryn Mortashed kmortashed93@gmail.com 936-577-8572 Beaches to Bayou Creations 23-034 Larriesha Legier rlarriesha@hotmail.com 225-439-1393 Fleur De Leigh Embroidery, LLC 23-035 Roxana Vazquez dreamcatcherboutique.rc@gmail.com 915-777-6985 Dream Catcher Boutique 23-036 Ariel Miller theenergizerbunnnies@gmail.com 910-303-9593 The Energizer Bunnies 23-038 Nina Morris ninarmorris@outlook.com 812-801-6997 The Toasted Tulip 23-039 Nicole Lipe lipeluffaco@gmail.com 618-521-1394 Lipe's Luffa Co. 23-040 Judy A. Herb judyallenherb@gmail.com 843-610-4318 Encore Piano Studio 23-041 Kathryn N. Robles kmacshbb@gmail.com 803-843-4245 K-Mac's 23-042 Candice H. Dumbrique madhatters7448@gmail.com 931-305-9387 Mad Hatters 23-043 Samantha M. Kraemer levelupnutrition2023@gmail.com 207-231-4134 Level Up Nutrition 23-044 Clara I. Archilla J.archilla860@gmail.com 337-397-2142 Clara Santos 23-045 Tia M. Wilson tmwilson117@yahoo.com 215-688-3508 Cakery & Sweets by Tia Monique 23-046 Krystal Payton rosiesresins@yahoo.com 803-415-7883 Rosie's Custom Tumblers 23-047 Stephanie L. Parker srobinson1781@gmail.com 843-260-4390 Parker Place Hair Studio 23-048 Cory T. Clark cory.t.clark.mil@army.mil 254-402-4237 Combat Threading 23-049 Brittany N. Hill bnhill317@aol.com 706-973-1843 Heathen Hill Creations 23-050 Naakiska Cherif naakiska_cummings@Yahoo.com 912-257-0301 Balanced Living Co. 23-051 Ava N. Jednak avajednak@gmail.com 915-228-6275 Sunshine Canvas Co. 23-052 Joanna Kirk jcheer2121@yahoo.com 757-749-9595 Crafty Butterflies 23-053 Amy N. Bruno lulabellewooddesigns@gmail.com 623-206-9337 Lulabelle Wood Designs 23-054 Candice M. Brooks waxreverie@gmail.com 502-542-2702 Wax Reverie 23-055 Kelsey Lousch alittleoffthetopgrooming@gmail.com 717-961-6841 A Little Off the Top Pet Grooming 23-056 Aaliyah N. Hill yummymummies823@gmail.com 402-43-9913 Yummy Mummies 23-057 Shonda S. Cooper shonda_stewart@outlook.com 270-696-3223 Boujee Creations 23-058 Gabriela Braxton gabbie_gee@yahoo.com 337-487-0950 Mama's Batter 24-001 Amanda Harris sirensoddities@gmail.com (812) 987-4905 Sirens Oddities 24-002 Erika Jackson kekascreations@outlook.com 425-802-8752 Keka's Creations 24-003 Keniya More keniyamoreno@gmail.com 480-398-6480 Enchanting Raven Co. 24-004 Jason P. Kohne jasonkohne@live.com 260-579-3279 KohneStedCustoms/InsectaCurisities 24-005 Gabriella A. Rader alohamacsmacarons@gmail.com 484-347-6742 Aloha Macs

博士瓦基尔·塔卡耶夫
11) Büchel, J., Mingard, C., Takhaveev, V., Reinert, PB, Keller, G., Kloter, T., Huber, SM, McKeague, M. 和 Sturla, SJ, 2023. 胶质母细胞瘤药物替莫唑胺的 O6-甲基鸟嘌呤单核苷酸分辨率基因组图谱。bioRxiv,2023.12.12.571283。正在《核酸研究》中审查。10) Mingard, C., Battey, JN, Takhaveev, V., Blatter, K., Hürlimann, V., Sierro, N., Ivanov, NV 和 Sturla, SJ, 2023. 通过吸烟的各个成分剖析癌症突变特征。化学毒理学研究,36(4),第714-123页。9)Jiang, Y., Mingard, C., Huber, SM, Takhaveev, V., McKeague, M., Kizaki, S., Schneider, M., Ziegler, N., Hurlimann, V., Hoeng, J., Sierro, N., Ivanov, NV 和 Sturla, SJ,2023. 人类基因组中烷基化的量化和映射揭示了突变特征的单核苷酸分辨率前体。ACS Central Science,9(3),第362-372页。 8) Takhaveev, V.、Özsezen, S.、Smith, EN、Zylstra, A.、Chaillet, ML、Chen, H.、Papagiannakis, A.、Milias- Argeitis, A. 和 Heinemann, M., 2023. 生物合成过程的时间分离是造成芽殖酵母细胞周期中代谢振荡的原因。《自然代谢》,5(2),第 294-313 页。7) Ortega, AD#、Takhaveev, V.#、Vedelaar, SR、Long, Y.、Mestre-Farràs, N.、Incarnato, D.、Ersoy, F.、Olsen, LF、Mayer, G. 和 Heinemann, M., 2021. 一种用于报告糖酵解通量的果糖-1,6-双磷酸盐合成 RNA 生物传感器。 Cell Chemical Biology, 28(11), pp.1554-1568. 6) Monteiro, F., Hubmann, G., Takhaveev, V., Vedelaar, SR, Norder, J., Hekelaar, J., Saldida, J., Litsios, A., Wijma, HJ, Schmidt, A. 和 Heinemann, M., 2019. 使用正交合成生物传感器测量单个酵母细胞中的糖酵解通量。分子系统生物学, 15(12), p.e9071。 5) Leupold, S., Hubmann, G., Litsios, A., Meinema, AC, Takhaveev, V., Papagiannakis, A., Niebel, B., Janssens, G., Siegel, D. 和 Heinemann, M., 2019. 酿酒酵母在其复制生命周期中经历不同的代谢阶段。Elife, 8, p.e41046。4) Takhaveev, V. 和 Heinemann, M., 2018. 克隆微生物种群中的代谢异质性。Current opinion in microbiology, 45, pp.30-38。 3) Filer, D., Thompson, MA, Takhaveev, V., Dobson, AJ, Kotronaki, I., Green, JW, Heinemann, M., Tullet, JM 和 Alic, N., 2017. RNA聚合酶III限制TORC1下游的寿命。《自然》,552(7684),第263-267页。2) Suplatov, D., Kirilin, E., Arbatsky, M., Takhaveev, V. 和 Švedas, V., 2014. pocketZebra:一种通过对不同蛋白质家族的生物信息学分析自动选择和分类亚家族特异性结合位点的网络服务器。《核酸研究》,42(W1),第W344-W349页。 1) Suplatov, D., Kirilin, E., Takhaveev, V. 和 Švedas, V., 2014. Zebra:用于对不同蛋白质家族进行生物信息学分析的网络服务器。《生物分子结构与动力学杂志》,32(11),第 1752-1758 页。研究资助

Priestia Flexa对木薯中氰化物的排毒
木薯(Manihot esculenta)是高于大米和玉米的热带碳水化合物食物的第三大来源。也称为Mandioca,Manioc,Yuca或Tapioca。这是许多热带和亚热带发展中国家,尤其是在西非的主要主食根作物。在90多个国家/地区成长,在全球范围内,它是人类饮食中第六个最重要的能源来源,并且是大米,糖和玉米/玉米之后的第四个能源供应商(Heuberger,Heuberger,2005年)。研究人员已经开发了几种木薯的加工方法,目的是降低其毒性,同时将高度易腐的根转换为可以被视为更稳定的产品的产品。发酵,阳光干燥,浸泡以及干燥或烘烤的过程已被报道为过程(Irtwange&Achimba,2009年)。两种不同类型的木薯是甜木薯(Manihot Dulcis)和苦木薯(Manihot esculenta)。苦木薯与高水平的氰化糖苷有关。甜木薯被认为没有太多的氰化物。在木薯的局部分类中,有些品种被视为“甜”(即无毒理)。这导致消费者对应用简单治疗的自满情绪,以在消耗块茎之前降低氰化物水平。因此,缺乏对氰化物中毒的潜在危险的认识,这是消耗生木薯块茎的原因(Cornelius,Robert,Gaymary,James&Sakurani,2019年)。在木薯中,主要的氰化糖苷是Linamarin。这是因为研究表明,在某些地区,尤其是在东非,甚至那些被认为是人类灾难的木薯品种也是如此(Mburu,Njue&Sauda,2011年)。因此,根据Osuntokun(1994)的长期消费少量氰化物会引起严重的健康问题,例如热带神经病。Alitubeera,Eyu,Benon,Alex&Bao-Ping(2019)报告说,2017年涉及乌干达98人的氰化物中毒爆发,其中发生了两起死亡案件。加工不足也会导致高氰化物的暴露,这会导致严重疾病(例如Jorgensen,Bak,Busk,Sorensen,Sorensen,Olsen,Puonti-Kaerlas&Moller,2005年)。这种抗营养素的存在通过木薯中的野马酶通过水解减少。已经采用了几种加工方法来降低木薯根的毒性,并同时将高度易腐的根转化为更稳定的产品。这些包括晒干,浸泡和发酵,然后干燥或烘烤(Irtwange&Achimba,2009)。传统育种者已经产生了具有低氰化物潜力的木薯品种,但它们并未成功提供完全没有氰化糖苷的木薯品种(Ngudi,Kuo&Lambien,2003)。也少量存在的是lotaustralin(甲基中胺)。也存在酶的Linamarase酶。Linamarin被Linamarase催化,将其迅速水解为葡萄糖和丙酮氰基羟化蛋白。它还将lotaustralin水解为相关的氰氢蛋白酶和葡萄糖。丙酮氰基氢蛋白在中性条件下分解为丙酮和氰化氢(食品标准澳大利亚新西兰,2005年)。在木薯被食用的一些热带国家中,很难分析木薯中氰化物的数量,因为执行测定程序所需的设施不容易获得,并且获得准确的分析方法是另一个困难领域。

Nan Hua,博士-Weizhou Zhang实验室 Seyedehalaleh(Alaleh)Anvar -Weizhou Zhang实验室
1。Sajid Khan, Janet Wiegand, Peiyi Zhang, Wanyi Hu, Dinesh Thummuri, Vivekananda Budamagunta, Nan Hua , Lingtao Jin, Carmen J Allegra, Scott E Kopetz, Maria Zajac-Kaye, Frederic J Kaye, Guangrong Zheng, Daohong.bcl- XL Protac DEGRADER DT2216与Sotorasib在KRAS G12C突变癌症的临床前模型中协同作用。PMID:35260176 2。Dongwen Lv, Pratik Pal, Xingui Liu, Yannan Jia, Dinesh Thummuri, Peiyi Zhang, Wanyi Hu, Jing Pei, Qi Zhang, Shuo Zhou, Sajid Khan, Xuan Zhang, Nan Hua , Qingping Yang, Sebastian Arango, Weizhou Zhang, Digant Nayak, Shaun K Olsen,Susan T Weintraub,Robert Hromas,Marina Konopleva,Yaxia Yuan,Guangrong Zheng,Daohong Zhou。开发具有改善抗白血病活性的BCL-XL和BCL-2双降解器。PMID:34824248 3。Pratik Pal,Dinesh Thummuri,Dongwen LV,Xingui liu,Peiyi Zhang,Wanyi Hu,Saikat K Poddar,Nan Hua,Sajid Khan,Yaxia Khan,Yaxia Yuan,Yaxia Yuan,Xuan Zhang,Xuan Zhang,Daohong Zhun Zhoun Zhoung,Guangangong Zheng。发现具有增强BCl-2抑制作用的新型Bcl-XL Protac降解器。PMID:34533954 4。Xu -Xu Zhuang,Xuan Zang,Zheng,Nan Hua,Yi Sun,Yu -Hui Hu,Ling He。聚丙醇减轻APP/PS1小鼠中的认知功能障碍和神经病理学。pmid:29468792 5。Xuan Zang,Zhao-Yan Cheng,Yi Sun,Nan Hua,Li-Hua Zhu,Ling He。多巴胺D1样受体激动剂SKF38393对Aβ1-42诱导的认知障碍的改善作用和潜在机制。PMID:28939187 6。Xuan Zang,Zhao-Yan Cheng,Yi Sun,Nan Hua,Li-Hua Zhu,Ling He。PMID:28939187 7。PMID:25858697 8。9。多巴胺D1样受体激动剂SKF38393对Aβ1-42诱导的认知障碍的改善作用和潜在机制。Yu-hang锣,Nan Hua,Xuan Zang,Tao Huang,Ling He。褪黑激素可以改善小鼠模型中Aβ1-42诱导的阿尔茨海默氏症的认知缺陷。Madison E Carelock,Rohan P Master,Myung-Chul Kim,Zeng Jin,Lei Wang,Chandra K Maharjan,Nan Hua,Umasankar de,Ryan Kolb,Yufeng Xiao,Yufeng Xiao,Daiqing Liao,Guangrong Zheng Zheng Zheng,Weizhou Zhun。针对癌症免疫疗法的细胞类型特异性功能靶向细胞内蛋白。Sajid Khan, Patrick Kellish, Nick Connis, Dinesh Thummuri, Janet Wiegand, Peiyi Zhang, Xuan Zhang, Vivekananda Budamagunta, Nan Hua , Yang Yang, Umasankar De, Lingtao Jin, Weizhou Zhang, Guangrong Zheng, Robert Hromas, Christine Hann, Maria Zajac-kaye,Frederic J Kaye,Daohong Zhou。与DT2216和AZD8055共同靶向BCl-XL和MCL-1,可以协同抑制小细胞肺癌的生长,而不会引起小鼠的靶向毒性。PMID:36588105。10。Jing Pei, Yufeng Xiao, Xingui Liu, Wanyi Hu, Amin Sobh, Yaxia Yuan, Shuo Zhou, Nan Hua , Samuel G Mackintosh, Xuan Zhang, Kari B Basso, Manasi Kamat, Qingping Yang, Jonathan D Licht, Guangrong Zheng, Daohong Zhou, Dongwen LV。哌拉金偶联物诱导靶向蛋白质降解。PMID:36750097。

缺氧诱导因子2α促进保护性TH2细胞...
Hypoxia-inducible factor 2 α promotes protective Th2 cell responses during intestinal 1 helminth infection 2 3 Jasmine C. Labuda 1 , Tayla M. Olsen 1,2 , Sheenam Verma 1 , Samantha Kimmel 1 , Thomas H. 4 Edwards 3 , Matthew J. Dufort 3 , Oliver J. Harrison 1,4 5 6 1 Center for Fundamental Immunology, Benaroya Research美国华盛顿州西雅图研究所。7 2分子和蜂窝生物学计划,美国华盛顿州西雅图市华盛顿大学。8 3美国华盛顿州西雅图市贝纳罗亚研究所系统免疫学中心。9 4美国华盛顿州华盛顿大学华盛顿大学免疫学系。10 11通信:oharrison@benaroyaresearch.org 12 13摘要:TH2细胞必须感知并适应组织环境,以提供保护性宿主14免疫和组织修复。在这里,我们检查了促进Th2细胞15分化和功能的机制。单细胞RNA-seq 16分析来自小肠道椎板椎板的CD4 + T细胞17揭示了基因EPAS1的高表达,编码了转录因子缺氧缺氧诱导的18因子2a(HIF2α)。在体外,即使在非极化条件下,暴露于缺氧或遗传HIF2α激活也促进了Th2细胞19分化。在小鼠中,CD4 + T细胞中的HIF2α激活20在没有感染的情况下促进了肠道Th2细胞的积累,而HIF2α缺陷21受损的CD4 + T细胞介导的宿主对肠舵感感染的免疫免疫。24 25简介:肠蠕虫感染是全球最普遍的慢性感染26。我们的发现22确定了缺氧,氧调节的转录因子缺氧诱导因子2α23(HIF2α)是小肠内Th2细胞分化和功能的关键调节剂。Helminth infections are often associated with polarized “type 2” immunity, including 27 activation and accumulation of T helper 2 (Th2) cells, type-2 innate lymphoid cells (ILC2), tissue 28 basophils and eosinophils, elevated serum immunoglobulin E (IgE), alternative activation of 29 macrophages and alterations of epithelial differentiation and mucus production that统称30重塑感染的解剖部位2。免疫事件和31个组织重塑的类似级联反应引发局部组织病理学发生在过敏性疾病中,包括过敏32哮喘3。33 34指导屏障组织中Th2细胞分化的机制尚不清楚。35然而,证据支持组织微环境在建立36保护性Th2细胞分化和功能中的指导性作用,这是由染色质访问性37和/或基因表达的变化提供的,在将Th2细胞从淋巴结到本塞质体38组织4,5的TH2细胞转运后的基因表达。组织警报蛋白,包括IL-25,IL-33和TSLP是在39个屏障组织中产生的关键因素,这些因素在Helminth 40感染6,7期间共同促进2型免疫力和Th2细胞反应。在41个屏障组织中影响Th2细胞功能的组织环境中其他提示的身份仍有待鉴定。42 43缺氧诱导因子(HIF)是介导细胞的关键转录因子,对缺氧8的有机体反应4。Consisting of 3 family members, (HIF1 α , HIF2 α and HIF3 α , 45 encoded by Hif1a , Epas1 and Hif3a, respectively), HIFs are post-translationally modified in an 46 oxygen-dependent enzymatic cascade that regulates their stability, nuclear translocation, 47 binding to hypoxia-response elements (HRE) and transcription of低氧诱导基因8。48在常氧条件下,HIF蛋白通过氧气在关键的脯氨酸残基上通过氧气-49依赖性丙酰羟化酶(PHD)酶羟基氧化。通过50