XiaoMi-AI文件搜索系统
World File Search SystemOncogene

简历 Silvia Giordano 个人信息 出生日期:...
研究主要课题 Silvia Giordano 在癌症领域有着丰富的研究经验,她主要研究信号转导,并逐渐从癌症基础研究转向转化肿瘤学。她的主要成就包括: • MET 基因编码的酪氨酸激酶的鉴定和生化/生物学特性:(i) 确定受体的结构并鉴定其在人体细胞中的组成性激活(Giordano,Nature,1989);(ii) 研究其生物合成和翻译后修饰(Giordano,Oncogene,1989)以及信号转导机制(Ponzetto,Cell,1994);(iii) 表征 MET 受体激活后促进的生物活性(Giordano,PNAS,1993);(iv) 记录原发性肿瘤中 MET 的过度表达和继发性病变中的基因扩增(Di Renzo,Clin. Cancer Res,1995); (v) 研究遗传性肾乳头状癌患者中发现的组成性活性 MET 突变形式的生化和生物学特性 (Giordano, FASEB, 2000; Michieli, Oncogene 1999; Bardelli, PNAS, 1998);(vi) 在具有 MET 组成性激活的肿瘤中发现 MET 成瘾 (Corso, Oncogene, 2008);(vii) 发现控制受体降解的新机制 (Petrelli, Nature, 2002; Foveau, Mol. Cell. Biol, 2009; Ancot, Traffic, 2012)。这些研究为发现该受体的结构和功能改变奠定了基础,有助于证明 MET 基因在不同类型人类肿瘤中的调控改变,并将 MET 确定为治疗靶点。 • 确定了不依赖配体的 MET 激活机制:由于与 Plexins(缺乏酪氨酸激酶活性的膜受体)相互作用而激活;该机制在癌细胞中的作用(Giordano,Nature Cell Biol.,2002;Barberis,Faseb J.,2004;Artigiani,EMBO. Rep.,2004;Conrotto,Oncogene,2004);Plexin 介导的 MET 激活的促血管生成作用(Conrotto,Blood,2005;Sierra,J. Exp. Med.,2008)。这些研究确定了 MET 激活的新模式,并暗示 MET 抑制可能对肿瘤细胞具有直接的抗肿瘤作用,对内皮细胞具有抗血管生成作用。 • 鉴定了肝细胞癌早期发展过程中涉及的分子病变(Kowalik, Hepatology, 2011;Petrelli, Oncogene, 2012 和 Hepatology 2014;Perra, J. Hepatology, 2014;Frau, Hepatology, 2015;Zavattari, Hepatology, 2015;Kowalik, Oncotarget, 2015,2016;Mattu, J Hepatology, 2016;Orru, Cancers, 2020;Kowalik, J Hepatol. 2020;Mattu, Cell Mol Gastroeterol Hepatol, 2022)。这些论文确定了基因和微小 RNA 在人类和实验性 HCC 发病中的作用,并强调了新的治疗靶点。 • 鉴定了针对酪氨酸激酶的靶向治疗的耐药机制:EGFR受体家族成员的激活以及MET和KRAS扩增在MET抑制剂耐药中的作用(Apicella,Oncogene,2016;Martin,Mol. Onc. 2014;Corso,Mol Cancer,2010;Cepero,Cancer Res,2010;Apicella,Cell Metabolism,2018;Migliore,EMBO Mol Med. 2018);MET扩增在EGFR抑制剂耐药中的作用(Bardelli,Cancer Discovery,2013);胆管癌对FGFR2抑制的耐药机制(Cristinziano,J. Hepatology,2021)。这些研究有助于预防耐药性的发生,对于MET驱动的获得性耐药的结肠癌患者和MET扩增的胃癌患者,可以为患者提供新的治疗选择。 • 建立胃癌患者来源的异种移植平台并确定新的分子靶点(Apicella,Oncogene,2017;Pietrantonio,Clin. Cancer Res,2018;Corso,Neoplasia,2018;Corso,Cancer Res,2019,2021;Ughetto,Gastric Cancer,2021,

MDT 讨论 - 2024 年阿斯利康肺脏峰会
ALK,间变性淋巴瘤激酶;BRAF,v-raf 鼠肉瘤病毒致癌基因同源物 B1;EGFR,表皮生长因子受体;ERBB2,v-erb-b2 禽红细胞白血病病毒致癌基因同源物 2;IHC,免疫组织化学;KRAS,Kirsten 大鼠肉瘤病毒致癌基因同源物;MET,间充质上皮转变因子受体;PD-L1,程序性死亡配体 1;ROS1,ROS 原癌基因 1;TTF-1,甲状腺转录因子 1;WT,野生型。

干细胞的分离和功能表征 -
• Homogenous starting material, continuous growth (Tiered Cell Bank concept: MCB/WCB) • Supply of a whole product life cycle (from pre-clinical to end of commercial life) • Virus-free cell line generation, only animal-free materials, Sartorius media in the bioreactor • Human Telomerase is not an Oncogene, no oncogenes used for “immortalization” • MSC/TERT display MSC-like morphology and电势

Atirmociclib (PF-07220060) 和 PF-07104091 是首批
参考文献: 1. Choi YJ, Anders L. 通过细胞周期蛋白 D 依赖性激酶进行信号传导。Oncogene。2014 年 4 月 10 日;33(15):1890-903。doi: 10.1038/onc.2013.137 2. VanArsdale 等人。针对细胞周期蛋白 D-CDK4/6 轴进行癌症治疗。Clin Cancer Res。2015 年 7 月 1 日;21(13):2905-10。doi: 10.1158/1078-0432.CCR-14-0816 3. Fassl 等人。CDK4 和 CDK6 激酶:从基础科学到癌症治疗。Science。2022 年 1 月 14 日;375(6577):eabc1495。 doi: 10.1126/science.abc1495
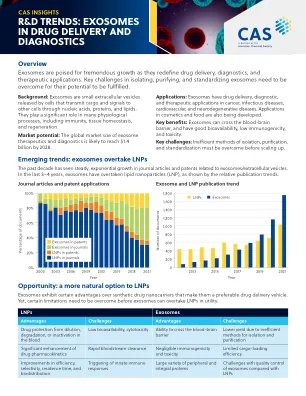
药物输送和诊断中的外泌体
缩写:ards =急性呼吸窘迫综合征; CTE =慢性创伤性脑病; ERG = V-ETS红细胞增生病毒E26癌基因同源物; exos =外泌体; LRKK2 =富含亮氨酸的重复激酶2;

STAT3信号传导的简短更新:当前的挑战和未来方向
与其他Stat家族成员共享序列和结构同源性,其细胞内功能有所不同,从而导致高度复杂的信号网络。例如,STAT1和STAT3具有很大的序列相似性,并且作为转录因子的行为,但是在许多生理条件下,它们受到相互调节,以在细胞增殖和凋亡细胞死亡中扮演相反的作用[3]。与STAT3形成鲜明对比的是癌基因,活化的STAT1充当肿瘤抑制器[3]。重要的是,STAT5与STAT3几乎没有序列相似性,也被认为是癌基因,主要是在造血恶性肿瘤中[4]。此外,STAT3激活失调通常与抗临床上可用的分子靶向剂的抗性相关[5]。这些功能使STAT3成为癌症治疗的吸引力,这促使靶向方法的发展[6-9]。

多种内分泌肿瘤2型2中有什么新功能?
ret(转染后重新排列)原癌基因在1985年被发现为癌基因。1 RET癌基因位于染色体10(10q.11.2)上的编码酪氨酸激酶家族的受体,这对于神经系统的发展以及源自神经crest的器官和组织的发展很重要。3种RET形式中的每一个都由3个不同的转录本编码。3个成绩单都有ER EXON 19;但是,随着外显子19的3'端的可变剪接,它们形成未剪接的外显子19,外显子20和外显子21。RET同工型具有9(RET9),51(RET51)和43(RET43)氨基酸C末端末端,由这些转录本编码。体内主要同工型为ret9和ret51,分别由1072和1114氨基酸组成。尽管在大多数组织中共表达,但这两种同工型具有不同的发育作用和基因表达谱,这意味着在细胞细胞接触途径调节中可能存在差异。2,3

IOV-COM-202 遗传修饰 Natarajan Guess,1 Anand Veerapthran。
*必须天真。†或≤4线如果≥2TKI。‡必须包括PD-1阻断抗体。§除了那些对靶向疗法敏感的已知癌基因驱动因素的患者。||如果BRAF突变为阳性,则可能已经收到Brafi/Meki。¶如果BRAF突变为阳性,则必须收到Brafi/Meki。#如果已知的癌基因驱动器突变,则必须接受有针对性的治疗。缩写:BRAFI,BRAF抑制剂; CPI,检查点抑制剂; ECOG,东部合作肿瘤学小组; FEV 1,在1秒内强迫呼气量; HNSCC,头颈部鳞状细胞癌; ICI,免疫检查点抑制剂; Meki,Mek抑制剂; NSCLC,非小细胞肺癌; PD-1,程序性细胞死亡蛋白1;恢复,实体瘤的反应评估标准; TIL,肿瘤浸润淋巴细胞; TKI,酪氨酸激酶抑制剂。

针对RET融合肺癌的靶向治疗
靶向突变或重排的癌基因驱动因素已成为晚期和复发性非细胞肺癌患者的标准公认治疗方法之一。ret位于人类10号染色体的长臂中,编码受体酪氨酸激酶蛋白,而RET融合阳性肺腺癌发生在1% - 2%的病例中。抑制RET致癌基因活性的多次蛋白酶抑制剂的临床试验,包括Cabozantinib,Vandetanib,Sorafenib和Lenvatinib的临床试验。最近,还已经开发了专门用于RET激酶的RET抑制剂,例如pralsetinib和selpercatinib,并且在先前的临床试验中研究了它们的效率(Blu-667和Loxo-292)。在这篇综述中,我们总结了多次激酶和选择性RET抑制剂的效果和不良事件,以及用于RET基因融合的各种诊断技术。在角度上,我们专注于RET融合阳性肺癌和未来发展的未解决问题。

使用靶向和全基因组测序对 SCLC 患者的循环游离 DNA 进行分析
SCLC 肿瘤的基因组分析已发现大量拷贝数变异 (CNA) 和高突变率。 4、5 肿瘤抑制基因肿瘤蛋白 p53 基因( TP53 )和视网膜母细胞瘤 1 基因( RB1 )几乎普遍失活和普遍丢失,转录调节因子(SRY-box 转录因子 2 基因 [ SOX2 ]、核因子 IB 基因 [ NFIB ]、v-myc 禽类粒细胞瘤病毒致癌基因肺癌衍生同源基因 [ MYCL1 ]、v-myc 禽类粒细胞瘤病毒致癌基因神经母细胞瘤衍生同源基因 [ MYCN ] 和 v-myc 禽类粒细胞瘤病毒致癌基因同源基因 [ MYC ])频繁扩增,Notch 和组蛋白乙酰转移酶基因(CREB 结合蛋白基因 [ CREBBP ] 和 E1A 结合蛋白 p300 基因 [ EP300 ])反复突变。 4-6 肿瘤活检,尤其是连续活检,对于 SCLC 来说是一项重大挑战,7,8