XiaoMi-AI文件搜索系统
World File Search SystemPMS2

OncoTrack 液体
ABL1、ABL2、AKT1、ALK、APC、AR、ARAF、ARID1A、ARID1B、ATM、ATR、ATRX、BAP1、BARD1、BRAF、BRCA1、BRCA2、BRIP1、C11orf65、CCND1、CDH1、CDK12、CDK4、CDKN2A、CDX2、CHEK1、CHEK2、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCL、FBXW7、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、FOXA1、FOXL2、GATA3、GNA11、GNAQ、GNAS、HNF1A、HRAS、IDH1、IDH2、INPP4B、 JAK1、JAK2、JAK3、KDM5C、KDM6A、KEAP1、KIT、KRAS、MAP2K1、MAP2K2、MAPK1、MET(外显子 14 跳跃)突变)、MLH1、MPL、MSH2、MSH6、MTOR、MUTYH、MYC、MYCN、MYD88、NF1、NF2、NOTCH1、NPM1、NRAS、NTRK1、NTRK3、PALB2、PBRM1、PDGFRA、PIK3CA、PMS2、POLD1、POLE、PPP2R2A、PTCH1、PTEN、PTPN11、RAD51B、RAD51C、RAD51D、RAD54L、RAF1、RB1、RET、RHEB、RHOA、RIT1、ROS1、SETD2、SF3B1、 SMAD4、SMARCB1、SMO、SPOP、SRC、STK11、TERT、TP53、TSC1、TSC2、VHL

Dragen TSO 500分析软件发行说明
•已知某些区域难以顺序。一个例子是TERT启动子区域。尽管可以在TERT启动子区域进行测序,但由于测序区域的GC含量富含GC,该位置可能会导致覆盖率较低。另一个示例区域是PMS2基因,该基因与伪基和读取可能无法正确对齐。通常,TSO 500面板旨在针对独特的区域,并且该软件在小型变体中说明了每个基因组位置的背景噪声。此设计旨在防止误报。分析性能的分析性能是评估整个面板的,而不是每个基因或外显子。但是,由于这些挑战,由于高背景噪声,产品清单中涵盖的某些区域被排除在分析之外。使用标志在VCF中识别所有排除的变体。此块列表包括以下基因:HLA-A,HLA-B,HLA-C,KMT2B,KMT2C,KMT2D,KMT2D,CHRY和VAF> 1%的位置发生在60个基线样本中的六个或更多。可以根据当地Illumina代表的要求获得排除站点的块列表。

FEP 2.04.02遗传性乳腺/卵巢癌综合征和其他高风险癌(BRCA1,BRCA2,PALB2)的生殖基因测试
2.04.126 - Germline Genetic Testing for Gene Variants Associated With Breast Cancer in Individuals at High Breast Cancer Risk (CHEK2, ATM, and BARD1) 2.04.128 - Genetic Testing for Fanconi Anemia 2.04.148 - Germline Genetic Testing for Pancreatic Cancer Susceptibility Genes (ATM, BRCA1, BRCA2, CDKN2A, EPCAM, MLH1, MSH2, MSH6, PALB2, PMS2, STK11, and TP53) 2.04.151 - Germline and Somatic Biomarker Testing (Including Liquid Biopsy) for Targeted Treatment and Immunotherapy in Breast Cancer 2.04.155 - Germline and Somatic Biomarker Testing (Including Liquid Biopsy) for Targeted Treatment and Immunotherapy in Prostate Cancer (BRCA1/2, Homologous Recombination Repair Gene Alterations, Microsatellite Instability/Mismatch Repair, Tumor Mutational Burden) 2.04.156 - Germline and Somatic Biomarker Testing (Including Liquid Biopsy) for Targeted Treatment and Immunotherapy in Ovarian Cancer (BRCA1, BRCA2, Homologous Recombination Deficiency, Tumor Mutational Burden, Microsatellite Instability/Mismatch Repair) 2.04.157 - Somatic Biomarker Testing for Immune Checkpoint抑制剂疗法(BRAF,MSI/MMR,PD -L1,TMB)2.04.93-使用下一代测序

tlr-2和
摘要遗传物质的稳定性和完整性对于维持和延续生活至关重要。人类基因组由三十亿对碱基组成,编码30,000-40,000个基因,并不断受到内源性反应性代谢产物,治疗药物和众多影响其完整性的环境诱变药物的攻击。因此,很明显,基因组的稳定性必须在连续监测之下。这是通过DNA修复机制实现的,DNA修复机制已开发出来去除或耐受DNA损伤和误差。在生物体中存在的DNA修复机制中,它们可以分为:i)基础切除修复(BER),ii)核苷酸切除(NER),iii)基本MALPASE(MMR)和IV)DNA修复,通过非同型末端(NHEJ)。对于这些机制正常工作,很明显,负责修复功能的蛋白质之间相互作用的重要性,以及对负责提到的机制的蛋白质正确位置的核进口调节。在负责调节核进口的机制中,由进口异二聚体α/β组成的经典途径是位移的主要机制之一。某些修复蛋白似乎仅与进口α(IMP)的某些同样蛋白相互作用,表明对修复过程的额外调节,但对这些蛋白质的核位置序列(NLS)的识别知之甚少。通过这些结果,阐明了包含NLSS KU80和FEN1的结构。这项工作特别涉及使用晶体学技术与蛋白质相关DNA修复的NLSS肽的IMP复合物的结构复合物的研究。进行了在其N末端部分截断的Musculus印象的表达和纯化,以及与DNA相关蛋白的NLS肽的IMP偶然化,对应于KU80,PMS2和MLH1蛋白质和MLH1蛋白质和BIPARTARTARTARTATTITE序列的单型序列。X射线衍射数据,并以2.1-2.38Å的分辨率进行处理。肽NLS KU80与NHEJ修复有关,与主连接位点上的IMPα相似,类似于SV40 T抗原的NLS(S 1)。已经与ber修复有关的NLS FEN1肽与Sitia S 1和次级位点(S 2)有关,证明是两部分序列。此外,仅具有10种废物的Fen1肽接头区域使与IMPα的联系更好,并且与具有11-12废物的肽的连接相比,与IMPα的连接更扩展,可能更有利的构象。在连接位点上的特定位置被确认为必不可少的,以及在这些区域中保守的残留物,表明这些位点中分子间相互作用的重要性。此信息表明这些蛋白质可以通过IMP-α独立运输到核心,而无需与有关修复的其他蛋白质形成复合物。关键字:进口α,核进口,NLS,射线晶体学-X,KU80,FEN1,PMS2,MLH1。

在炎症和氧化应激微卫星机构中的作用
微粒细胞炎不稳定是一种遗传现象,其特征在于重复的核苷酸序列被称为微卫星。这种不稳定可能是由于DNA修复基因(例如MLH1,MSH2,MSH6和PMS2基因)的缺陷而发生的。慢性炎症与结直肠癌的发展有关。微卫星不稳定性基因参与调节炎症反应,并可能影响肿瘤进展。研究表明,胶体肿瘤中微卫星不稳定性的存在与更大的免疫细胞浸润有关,例如T淋巴细胞,巨噬细胞和中性粒细胞,这可能会调节肿瘤微环境中的炎症反应。氧化应激的特征在于氧气活性物种的产生与生物体的抗氧化能力之间的不平衡,并在癌变中起重要作用。微卫星不稳定性基因可以影响对氧化应激的反应,从而影响肿瘤细胞处理氧化损伤并促进细胞存活的能力。这项工作的目的是了解大肠癌中微卫星不稳定性涉及的基因,以及它们如何对疾病的发展做出贡献,与炎症过程和肿瘤细胞中氧化应激有关。这是有必要理解大肠癌患者的微囊炎,炎症和氧化应激之间的相互联系的合理性。关键词:微炎症的不稳定性;结直肠癌;炎;氧化应激。

人工智能生成的小活页夹改进了主要编辑
引物编辑 2 (PE2) 系统包含一个切口酶 Cas9,该切口酶与逆转录酶融合,利用引物编辑向导 RNA (pegRNA) 在目标基因组位点引入所需突变。然而,PE 效率受到错配修复 (MMR) 的限制,错配修复会切除包含所需编辑的 DNA 链。因此,通过显性负 MLH1 (MLH1dn) 的瞬时表达抑制 MMR 复合物的关键成分,PE 效率比 PE2 提高约 7.7 倍,从而生成 PE4。在此,通过利用生成人工智能 (AI) 技术 RFdiffusion 和 AlphaFold 3,我们最终生成了一种从头 MLH1 小结合物(称为 MLH1-SB),它与 MLH1 和 PMS2 的二聚体界面结合,以破坏关键 MMR 成分的形成。MLH1-SB 的尺寸很小(82 个氨基酸),因此可以通过 2A 系统将其整合到预先存在的 PE 架构中,从而创建一个新颖的 PE-SB 平台。结果,通过将 MLH1-SB 整合到 PE7 中,我们开发了一种改进的 PE 架构,称为 PE7-SB,它表现出迄今为止最高的 PE 效率(在 HeLa 细胞中是 PE2 的 29.4 倍,是 PE7 的 2.4 倍),这表明生成式 AI 技术将促进基因组编辑工具的改进。
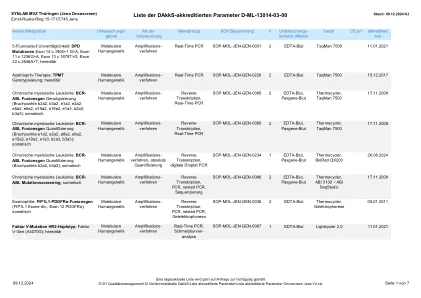
认可参数列表 Oncoscreen Jena v5.xlsx
Oncomine Comprehensive Assay v3 DNA 组:AKT1、AKT2、AKT3、ALK、AR、ARAF、ARID1A、ATM、ATR、ATRX、AXL、BAP1、BRAF、BRCA1、BRCA2、BTK、CBL、CCND1、CCND2、CCND3、CCNE1、CDK12、CDK2、CDK4、CDK6、CDKN1B、CDKN2A、CDKN2B、CHEK1、CHEK2、CREBBP、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCA、FANCD2、FANCI、FBXW7、FGF19、FGF3、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、 FOXL2、GATA2、GNA11、GNAQ、GNAS、H3-3A、HIST1H1E、HNF1A、HRAS、IDH1、IDH2、IGF1R、JAK1、JAK2、JAK3、KDR、KIT、KNSTRN、KRAS、MAGOH、MAP2K1、MAP2K2、MAP2K4、MAPK1、MAX、MDM2、 MDM4、MED12、MET、MLH1、MRE11A、MSH2、MSH6、MTOR、MYC、MYCL、MYCN、MYD88、NBN、NF1、NF2、NFE2L2、NOTCH1、NOTCH2、NOTCH3、NRAS、NTRK1、NTRK2、NTRK3、PALB2、PDGFRA、PDGFRB、PIK3CA、 PIK3CB, PIK3R1、PMS2、POLE、PPARG、PPP2R1A、PTCH1、PTEN、PTPN11、RAC1、RAD50、RAD51、RAD51B、RAD51C、RAD51D、RAF1、RB1、RET、RHEB、RHOA、RICTOR、RNF43、ROS1、SETD2、SF3B1、SLX4、SMAD4、SMARCA4、SMARCB1、SMO、SPOP、SRC、STAT3、STK11、TERT、TOP1、TP53、TSC1、TSC2、U2AF1、XPO1

BRD4 抑制会损害 DNA 错配修复,诱导错配修复突变特征,并导致 MMR 功能正常的肿瘤对免疫检查点阻断的治疗脆弱性
摘要 背景 错配修复缺陷 (dMMR) 是免疫检查点阻断 (ICB) 反应的一个公认的生物标志物。将 MMR 熟练 (pMMR) 转化为 dMMR 表型以使肿瘤对 ICB 敏感的策略受到高度追捧。含溴结构域 4 (BRD4) 抑制和 ICB 的结合提供了有希望的抗肿瘤作用。然而,其潜在机制仍然未知。在这里,我们发现 BRD4 抑制会在癌症中诱导持续的 dMMR 表型。方法我们通过对癌症基因组图谱和临床蛋白质组肿瘤分析联盟数据进行生物信息学分析以及对卵巢癌标本的免疫组织化学 (IHC) 评分进行统计分析,证实了 BRD4 与错配修复 (MMR) 之间的相关性。通过定量逆转录 PCR、蛋白质印迹和 IHC 测量 MMR 基因 (MLH1、MSH2、MSH6、PMS2)。通过全外显子组测序、RNA 测序、MMR 检测和次黄嘌呤-鸟嘌呤磷酸核糖转移酶基因突变检测确认 MMR 状态。在体内和体外诱导 BRD4i AZD5153 耐药模型。通过细胞系之间的染色质免疫沉淀和来自 Cistrome 数据浏览器的数据研究了 BRD4 对 MMR 基因转录的影响。在体内证明了对 ICB 的治疗反应。通过流式细胞术测量了肿瘤免疫微环境标志物,例如 CD4、CD8、TIM-3、FOXP3。结果我们在转录和翻译方面确定了 BRD4 和 MMR 基因之间的正相关性。此外,BRD4 转录抑制会降低 MMR 基因表达,导致 dMMR 状态和突变负荷升高。此外,长期暴露于 AZD5153 可在体内和体外促进持久的 dMMR 特征,增强肿瘤的免疫原性,并且尽管获得了耐药性,但仍增加了对 α - 程序性死亡配体-1 疗法的敏感性。

从 70 万名生物库参与者中洞察 DNA 重复扩增的原因和后果
从 700,000 名生物库参与者的数据中深入了解 DNA 重复扩增的原因和后果 Margaux LA Hujoel 1,2,3,*、Robert E. Handsaker 3,4,5、Nolan Kamitaki 1,2,3,6、Ronen E. Mukamel 1,2,3、Simone Rubinacci 1,2,3、Pier F. Palamara 7,8、Steven A. McCarroll 3,4,5、Po-Ru Loh 1,2,3,* 1 美国马萨诸塞州波士顿布莱根妇女医院和哈佛医学院医学系遗传学分部 2 美国马萨诸塞州波士顿布莱根妇女医院和哈佛医学院数据科学中心 3 美国马萨诸塞州剑桥麻省理工学院和哈佛大学布罗德研究所医学和群体遗传学项目 4 美国马萨诸塞州波士顿麻省理工学院和哈佛大学布罗德研究所斯坦利精神病学研究中心。 5 美国马萨诸塞州波士顿哈佛医学院遗传学系。 6 美国马萨诸塞州波士顿哈佛医学院生物医学信息学系 7 英国牛津大学统计学系 8 英国牛津大学人类遗传学中心* 通讯作者:mhujoel@broadinstitute.org (MLAH),poruloh@broadinstitute.org (P.- RL) 摘要串联 DNA 重复的扩增和收缩是人类群体和人类组织中遗传变异的来源:一些扩增的重复会导致遗传疾病,一些还会造成体细胞不稳定。我们分析了来自英国生物银行和“我们所有人”研究计划中 700,000 多名参与者的血细胞的 DNA 序列数据,并开发了新的计算方法来识别、测量和学习 15 个高度多态性的 CAG 重复位点的 DNA 重复不稳定性。我们发现,即使对于相同长度的等位基因,这 15 个基因座的扩张和收缩率也差异很大;不同基因座的重复序列在生殖系和血液中也表现出差异很大的相对突变倾向。TCF4 重复序列的高度体细胞不稳定性使得全基因组关联分析成为可能,该分析确定了七个基因座,在这些基因座上,遗传变异会调节血细胞中的 TCF4 重复不稳定性。其中三个相关基因座所含基因( MSH3 、 FAN1 和 PMS2 )也会调节亨廷顿氏病的发病年龄以及血液中 HTT 重复的体细胞不稳定性;然而,特定的遗传变异及其效应(不稳定性增加或减少)似乎是组织特异性和重复特异性的,这表明不同组织中的体细胞突变(或同一组织中不同重复的体细胞突变)是独立进行的,并受截然不同的遗传变异的控制。其他修饰基因位点包括 DNA 损伤反应基因 ATAD5 和 GADD45A。分析 DNA 重复扩增并结合临床数据显示,谷氨酰胺酶 (GLS) 基因 5' UTR 中的遗传重复与 5 期慢性肾脏疾病 (OR=14.0 [5.7–34.3]) 和肝脏疾病 (OR=3.0 [1.5–5.9]) 相关。这些结果和其他结果都指出了人类群体和整个人类生命周期中 DNA 重复的动态。

摘要
RNA/DNA结合蛋白TDP43调节DNA不匹配修复基因1具有对基因组稳定性的影响2 3 Vincent E. Porpasek 1,2,Albino Bacolla 3,Albino Bacolla 3,Suganya Rangaswamy。1,Joy Mitra 1,Manohar 4 Kodavati 1,Issa O. Yusuf 4,Vikas H. Malojirao 1,Velmarini Vasquez 1,Gavin W. Britz 1,5,Guo-Min 5,Guo-Min 5 Li 6,li 6,Zuoshang Xu 4,Zuoshang Xu 4,Zuoshang Xu 4,Sankar Xu 4,Sankar Mitra 1,Sankar Mitra 1,Sankar Mitra 1,sankar M.Garrph M. Garrar and A. Hegde 1,8* 7 8 1神经循环中心的DNA修复研究部,美国德克萨斯州休斯敦市休斯顿市卫理公会研究所9神经外科977030。10 2美国德克萨斯州A&M大学医学院,美国德克萨斯州77843,美国。11 3分子和细胞肿瘤学系,癌症生物学系,德克萨斯大学医学博士12安德森癌症中心,休斯敦,德克萨斯州休斯敦,美国德克萨斯州77030,美国13 4 4 4 MASSACHUSETTS CHAN 14医学院的生物化学和分子生物技术系,MA约克,纽约10065,美国。17 6德克萨斯大学西南医学中心的辐射肿瘤学系,达拉斯,18 TX 75390,美国。19 7纽约州立大学宾厄姆顿大学生物科学系,纽约州宾厄姆顿20号,13902。21 8美国纽约市威尔·康奈尔医学院神经科学系,美国纽约10065,美国。22 23 *作者应向谁解决。24 25摘要26焦油DNA结合蛋白43(TDP43)越来越认可其参与27种神经退行性疾病,尤其是肌萎缩性侧面硬化症(ALS)和额叶28痴呆症(FTD)。TDP43蛋白质病,其特征在于核输出失调和29个细胞质聚集,并且与神经元中30个核功能和基因组不稳定的丧失有关。基于与TDP43 31病理与DNA双链断裂(DSB)的先前证据建立,本研究确定了在DNA不匹配修复(MMR)途径中32 TDP43的新调节作用。我们证明了TDP43的耗竭或33过表达会影响关键MMR基因的表达,包括MLH1,MSH6、34 MSH2,MSH3和PMS2。具体而言,TDP43通过替代剪接和转录本稳定性调节MLH1和MSH6 35蛋白的表达。这些发现在ALS 36小鼠模型,患者衍生的神经祖细胞和ALS 37例患者的尸体型脑组织中得到了验证。此外,MMR耗竭显示出神经元细胞中TDP43诱导的DNA损伤38的部分营救。TCGA癌症数据库的生物信息学分析揭示了TDP43与MMR基因表达式之间的显着39相关性与40种癌症亚型的突变负担之间的相关性显着。这些结果共同建立了TDP43作为MMR 41途径的关键调节剂,对理解基因组不稳定性42潜在的神经退行性和肿瘤性疾病具有广泛的影响。43