XiaoMi-AI文件搜索系统
World File Search SystemPOLD

RADA和DNA聚合酶在重组中的作用 - ... 对甲基甲基蛋白抗甲氧西林和甲氧西林敏感的金黄色葡萄球菌ST398菌株的基因组分析 氯化的双苯基转化,过氧化物酶和氧化酶活性的真菌和细菌从历史污染部位分离出来 在28nm n-mosfets中,氧化物分解事件的位置专用于RF应用程序 hi-ness:基于细菌核苷相关蛋白的遗传编码的DNA标记家族
摘要:在生命的三个领域中,同源重组(HR)的过程在修复双链DNA断裂和重新开始停滞的复制叉中起着核心作用。奇怪的是,参与人力资源过程的主要蛋白质参与者似乎对于高素化的古细菌提出了有关人力资源在极端条件下的古细菌中的复制和修复策略中的作用的有趣问题。该过程的一个关键参与者是重组酶RADA,它允许同源链搜索,并提供了遵循DNA合成并恢复遗传信息所需的DNA底物。DNA聚合酶在古细菌中尚不清楚链交换步骤后的操作。使用Abyssi Abyssi蛋白的工作,在这里我们表明,DNA聚合酶,家庭-B聚合酶(POLB)和家族-D聚合酶(POLD)都可以负责处理RADA介导的重组中间体。我们的结果还表明,与POLB相比,POLD的效果要少得多,以扩展位移环(D-Loop)底物处的入侵DNA。这些观察结果与先前对热圆菌物种获得的遗传分析相吻合,表明POLB主要参与DNA修复,而不是必不可少的,这可能是因为Pold可以接管其他伴侣。
●我们的结果表明IPS是可推广的多...
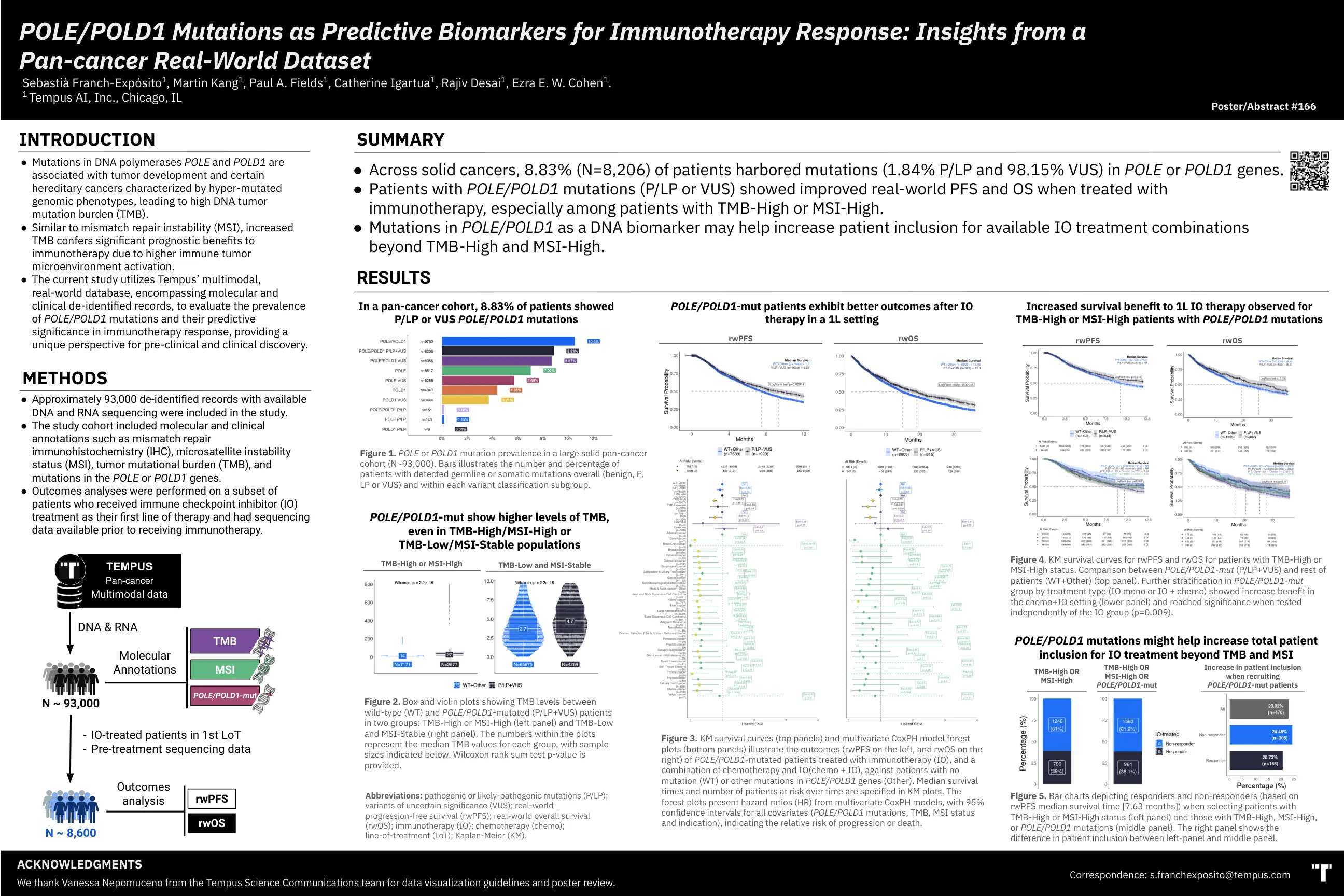
图3。km存活曲线(顶部面板)和多元Coxph森林图(底部面板)说明了POL/POLD 1的左侧的RWPF(左侧RWPF,右侧RWOS)的结果(RWOS),用免疫疗法(IO)治疗的患者(IO)以及与化学疗法和IO + IO + IO + IO + IO + IO + IO(IO)组合的结局(左侧),并与IO + IO + IO(IO)组合进行了突变(基因(其他)。在KM图中指定了随着时间的流逝的中位生存时间和处于危险中的患者人数。森林图具有多元COXPH模型的危险比(HR),所有协变量(POL/POLD1突变,TMB,MSI状态和指示)的置信间隔为95%,表明相对的进展或死亡风险。

结交朋友:新生儿免疫系统对共生体的积极选择和病原体的拒绝
北极陆地生态系统目前存储在地球高纬度地区的最大碳。在过去30年中,这些区域的温度水平的上升速度是全球平均水平的两倍,为每十年0.6℃(Cohen等,2014; Schuur等,2015)。这是一种强大的现象,称为北极扩增(Fengmin等,2019)。土壤微生物在将碳化合物转化为有机或无机化合物中起着重要作用,由于变暖,它们的代谢率提高。当微生物分解有机碳时,它们会释放温室气体(GHG),例如二氧化碳(CO 2),一氧化二氮(N 2 O)和甲烷(CH 4),导致全球气候变化(Mehmood等人,2020年,2020年; Marushchak等人,2021年)。在过去的800,000年中,大气二氧化碳,N2O和CH4的水平显着增加。CO 2的目前水平为390.5份百万分之390.5份,n 2 O的零件为390.5份(ppb),CH 4分别为1,803.2 ppb,这些水平分别为40、20、20和150%,比工业时代之前(Tian et et an e an and an an and an and and an and and and and and and and and and and and and and and and and and and and and and and。ch 4,仅次于CO 2之后的第二大最重要的温室气体,占自工业前时代以来变暖剂的人为辐射强迫的20%。此外,CH 4的温室作用是100年内CO 2的28倍(Tian等,2016; Ganesan等,2019; Hui等,2020)。在2000年至2017年之间的生物地球化学模型和大气反转估计,CH 4排放量为15至50 tg/yr(Saunois等,2016,2020)。在2000年至2017年之间的生物地球化学模型和大气反转估计,CH 4排放量为15至50 tg/yr(Saunois等,2016,2020)。由于北极扩增,全球气候变化将导致北极土壤变暖和CH 4排放。然而,尚未发现变暖对CH 4释放的影响,从而导致气候变化。微生物代谢过程长期以来一直是对气候变化的关键驱动因素和反应者(Singh等,2010)。根据研究发现,不同的土壤微生物通过与微生物组成相关的不同代谢途径产生温室气体,从而提高了对温室气体排放的理解。例如,大多数土壤微生物通过分解和异养呼吸对CO 2排放产生了巨大贡献(Watts等,2021)。类似于CO 2排放,生物CH 4的排放受土壤微生物甲烷生成和CH 4氧化的控制,来自土壤,湖泊和其他陆地陆地,尤其是北极土壤(Nazaries等,2013; Tveit et al。微生物甲烷生成是一组厌氧甲烷古细菌进行的过程(Song等,2021)。虽然其他微生物可以分解CH 4,从而减少CH 4向大气中的释放,但微生物甲烷发生对全球CH 4排放造成了很大的贡献,并且了解其对变暖时间的反应至关重要,这对于预测有效的温室气体和气候变化之间的反馈(Lee等人,2012年; Chen等,2020年)。此外,预计在按年来衡量的长期变暖的情况下,微生物组成将发生变化(Deslippe等,2012; Pold等,2021; Zosso等,2021; Rijkers等,2022; Zhou等,2023)。同时,生物CH 4排放也是由于长期微生物发酵而变暖引起的(Altshuler等,2019; Hui等,2020; Zhang等,2021)。但是,气候变化是一个过程